 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Actinomycetes, Vol. 10, 2000, pp 1-6 Fatty Acid Composition of Streptomyces hygroscopicus Populations V. GESHEVA and R. RACHEV Institute of Microbiology, Bulgarian Academy of Sciences, Sofia 1113 Bulgaria Code Number: AC00001 ABSTRACT. The composition of total fatty acids of Streptomyces hygroscopicus, strain 111-81 and of its spontaneous mutants was analysed by capillary gas chromatography. Parent culture and most variants, white and oligosporic showed similar fatty acid profiles and antibiotic metabolites. Asporogenic and Nocardia-like variants were characterised by different antibiotic spectra and by shifts in fatty acid ratios compared with the parent strain. Results are discussed with reference to antibiotic biosynthesis. Streptomycetes are extremely variable microorganisms with regard to aerial mycelium formation, sporulation, production of antibiotics and other bioactive metabolites (Rodionova & Danilenko, 1988; Leblond et al., 1991; Schneider et al., 1993; Marins et al., 1994; Dou et al., 1995; Qin et al., 1998). Most authors have described variability induced by mutagens (Ogura et al., 1986; Leblond et al., 1989, 1991; Marins et al., 1994; Golet et al., 1995; Vikhanski & Bartoshevich, 1996) and only few (Sokolova et al., 1987; Vesselinova & Gesheva, 1991; Gesheva & Gesheva, 1993). The Streptomyces hygroscopicus population has been shown (Gesheva & Gesheva, 1993) to consist of five morphological and cultural spontaneous variants. Fatty profiles of these strains have been analysed and compared with those of the parent culture. MATERIALS and METHODS Organisms and cultivation. S. hygroscopicus, strain 111-81, and some spontaneous variants were maintained on Mineral Agar No. 1, while asporogenic and Nocardia-like strains were kept on Maize Extract Agar No. 6 (Gesheva & Gesheva, 1993) Strains were grown as submerged cultures at 28°C in two stages, in 500 ml Erlenmeyer flasks (50 ml of medium) on a rotary shaker (220 rpm) to obtain good growth and then they were transferred to a fresh medium and harvested at the mid-exponential phase. Medium composition (pH 7.2) was as follows (g/l): glucose, 30; KNO3, 1; K2HPO4, 0.5; NaCl, 0.5; CaCO3, 1. Antibiotic assay. Cultures were processed according to Gesheva & Gesheva (1993). Antibiotic activity was determined by the agar diffusion method against Candida utilis as test culture and by thin-layer chromatography (TLC) according to Gesheva et al. (1994), using the non-polyenic macrolide antibiotic AK-111-81 as standard. Fatty acid analysis. The mycelium was harvested by centrifugation and then treated according to Gesheva et al. (1997). Gas chromatography was carried out in a C. Erba Science 4300 chromatograph under the following operating conditions: WCOT column DB5 (Varian) (25m x 0.22mm x 0.2µm) and Omegawax 250 (Supelco) (30m x 0.22mm x 0.2µm); initial temperature 90°C (5 min); program rate 5°C/min; final temperature 220°C (5 min); injection and detection temperatures 240°C; detector FID. Methyl esters and fatty acids were identified by comparison of retention time with standards (Polyscience, Co., USA). For each strain triplicate samples from three different cultures were tested. Attempts to separate C18:1 from C18:2 fatty acids with the WCOT column were unsuccessful; and the Omegawax column was therefore used. Table 1. Fatty acid composition of S. hygroscopicus, strain 111-81, and of its variants (ai: anteiso acid; i: iso acid; sc: saturated straight-chain acid; u: unsaturated acid; x: unidentified; A: asporogenic; M: main; NL: Nocardia-like; O: oligosporic; P: parent; W: white)
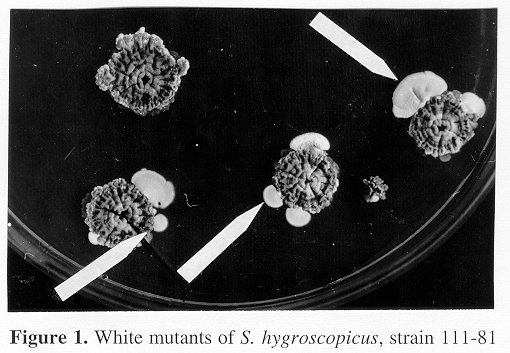
Figure 1. White mutants of S. hygroscopicus, strain 111-81 RESULTS and DISCUSSION The fatty acid composition of strain 111-81 and of its spontaneous variants is shown in Table 1. Main variants (Gesheva & Gesheva, 1993) are the most frequent (86.6%), they resembles the parent strain showing abundant sporulation and spiral sporophores. The aerial mycelium (a.m.) shows various shades of grey, turning black after lysis on some media. White strains (Fig. 1) are characterised by the colour of the a.m., which in some parts of the colony is sterile. Asporogenic cultures (0.53%) show no a.m. or just traces. Primitive open spirals are formed sometimes by White and Asporogenic cultures. Oligosporic cultures (4.55%) show a.m. which however does not uniformly cover the whole colony surface. Nocardia-like variants are rare (0.02%), the colonies are irregular in shape and have a soft consistency, the a.m. is scarce or absent, the substrate mycelium fragments and is yellow to ochre in colour. C16:0, C18:1w9 and C18:2 are the most abundant fatty acids in the parent culture (111-81) and in all the variant strains investigated. The composition of fatty acids is an important feature of the cell membrane function. Straight-chain saturated fatty acids are approximately equivalent in all the cultures with the exception of the Nocardia-like variant. This organism produces trace amounts of both IM-111-81 (non-polyenic macrolide) and AK-111-81 (polyether) antibiotic complexes on a soy medium (Gesheva & Gesheva, 1993). The parent culture (111-81) forms in addition other antibiotics such as azalomycin B (Gesheva et al., 1994) (Table 2). Table 2. Antibiotics produced by S. hygroscopicus, strain 111-81 and its variants on a synthetic medium (AZ: azalomycin B; tr: traces; +: positive; -: negative)
The fatty acid profile of the Nocardia-like variant is different from that of the parent strain. The low proportion of straight-chain saturated fatty acids is due perhaps to a different level of enzymes, such as crotonyl CoA reductase, a key enzyme in their biosynthesis in streptomycetes (Reynolds, 1995). According to Reynolds (1998) enzymes such as 3-ketoacylsynthase III regulate the formation of straight and branched long-chain fatty acids in streptomycetes and in E. coli. The Asporogenic variant shows some shifts in fatty acid profiles (C15:1, C20:0) and a higher proportion of iso-branched acid in comparison to the other strains. These differences may be associated with the impaired ability of the organism to differentiate. The antibiotic spectrum is also different. According to some authors (Arima et al., 1973; Grafe et al., 1982; Vancura et al., 1987; David et al., 1992) antibiotic biosynthesis in streptomycetes is closely associated with the structure and the function of the cell membrane. Differences in fatty acid composition influence membrane functions and in particular membrane permeability. Similar proportions of straight-chain saturated and insaturated fatty acid indicate a close relationship between the variants and also a common ancestor. However the variants are characterised by distinct antibiotic spectra. In our opinion the alteration of fatty acid ratios, affecting cell membrane properties, could consequently favour the biosynthesis of some antibiotics. REFERENCES Arima, K., H. Okazaki, H. Hono, K. Yamada & T. Beppu (1973). Effect of exogenous fatty acids on the cellular fatty acid composition and neomycin formation in a mutant strain of Streptomyces fradiae. Agr. Biol. Chem., 37: 2313- 2317 David, L., H. Loutheiller, D. Bauchart, S. Ausboiron & J. Asselineau (1992). Effects of exogenous methyloleate on the biosynthesis of nigericin, a polyether carboxylic antibiotic by Streptomyces hygroscopicus NRRL B-1865. Biosci. Biotechnol. Biochem., 56: 330 Dou, K, M.C. Saeki, Y. Ono & S. Ogata (1995). Plasmid formation and its relation to the formation of spontaneously developing pocks in Streptomyces azureus ATCC 14921. J. Appl. Bacteriol., 79: 257- 263 Gesheva,V. & R. Gesheva (1993). Structure of the Streptomyces hygroscopicus 111-81 population and characteristics of its variants. Actinomycetes, 4: 65- 72 Gesheva, V., V. Ivanova & R. Gesheva (1994). Biological characteristics and antibiotic production of Streptomyces hygroscopicus strains. Actinomycetes, 5: 57- 63 Gesheva, V., R. Rachev & S. Bojkova (1997). Fatty acid composition of Streptomyces hygroscopicus strains producing antibiotics. Lett. Appl. Microbiol., 24: 109- 112 Golets, L.M., I.I. Basiliya, A.I. Mazepa & V.A. Fedorenko (1995). Isolation of Streptomyces kanamyceticus mutants with impaired biosynthesis of kanamycins and investigations of their properties. Antib. Chemotherapy, 40: 3-7 Grafe U. , M. Roth & D. Krebs (1982). Effect of L-valine and L- isoleucine o fatty acid composition of Streptomyces hygroscopicus and Streptomyces griseus. Z. Allg. Microbiol., 22: 595- 599 Leblond, P., P. Demuyter, L. Mouter, M. Laakel & J.M. Simonet (1989). Hypervariability, a new phenomenon of genetic instability, related to DNA amplification in Streptomyces ambofaciens. J. Bacteriol., 171: 419- 423 Leblond, P., P. Demuyter, J.M. Simonet & B. Decaris (1991). Genetic instability and associated genome plasticity in Streptomyces ambofaciens: Pulsed field gel electrophoresis evidence for large DNA alterations in a limited genomic region. J. Bacteriol., 173: 4229- 4233 Marins, M., S. O. Costa & G. Padilla G. (1994). Effects of genotoxic agents on his+ revertants and survival of spores of Streptomyces aureofaciens. World J. Microbiol. Biotechnol., 10: 560- 562 Ogura M., T. Tanaka, K. Furihata, A. Shimazu & O. Otake (1986). Induction of antibiotic production with ethidium bromide in Streptomyces hygroscopicus. J. Antibiot., 39: 1443- 1450 Reynolds, K. A. (1995). Butyrate metabolism in Streptomyces. Abstr. SIM Ann. Meeting, San Jose, CA, 67 Reynolds, K. A. (1998). Working at the surface of fatty acids and polyketide biosynthesis. Abstr. VIII Int. Symp. Gen. Ind. Microorg., Jerusalem, Israel, 34 Rodoinova, I.I.& V.N. Danilenko (1988). Factors of Streptomyces genetic instability and their use in selection practice. Antib. Chemotherapy, 33: 177- 179 Qin, Z., X. Zhou, Q. Zhou, H. Chen, D. Hopwood, T. Kieser & Z. Deng (1998). Strain improvement by genetic manipulation for the overproduction of several chemically unrelated antifungal antibiotics which are used for plant protection. Abstr. VIII Int. Symp. Gen. Ind. Microorg., Jerusalem, Israel, 50 Schneider, D;, B. Aigle, P. Leblond, J.M. Simonet & D. Decaris (1993). Analysis of genome instability in Streptomyces ambofaciens. J. gen. Microbiol., 139: 2559- 2567 Sokolova, Z.G., K.A. Vinogradova & A.N. Polin (1987). Polymorphism of Streptomyces olivocinereus producer of geliomycin. Antib. Medic. Biotechnol., 32: 15- 19 Vancura, A., T. Rezanka, V. Marsalek, V. Kristan & G. Barasova (1987). Fatty acids and production of tylosin compounds in Streptomyces fradiae. J. Bas. Microbiol., 27: 167- 171 Vesselinova, N. & R. Gesheva (1991). Variability of Streptomyces spectabilis 1000. Actinomycetes, 2: 13- 17 Vikhansky, Y.D. & Y.E. Bartoshevich (1996). A targeted selection of the producer of omomycin. Biotechnologiya, 7: 23- 25. Copyright 2000 C.E.T.A., The International Centre for Theoretical and Applied Ecology, Gorizia The following images related to this document are available:Photo images[ac00001a.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||
{kind=link}