 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
A Method for Separating Aerial and Substrate Mycelia in Streptomycetes W.NEUMEIER (1) and E.KUSTER (2) (1) Medizinisches Zentrum fur Hygiene und Medizinische Mikrobiologie mit Medizinaluntersuchungsamt, Institut fur Umwelthygiene und Krankenhaushygiene, PhilippsUniversitat, Marburg and (2) Institut fur Mikrobiologie und Landeskultur, Justus-LiebigUniversitat, Giessen, BRD
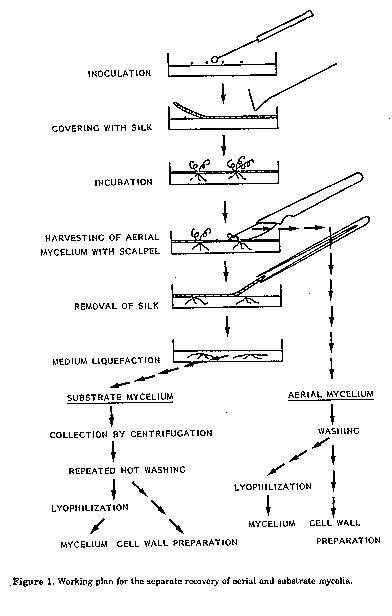
ABSTRACT. The separation of aerial and substrate hyphae is difficult because of the small height of the mycelia and biochemical differences are therefore little known. In the method described aerial hyphae of streptomycetes grew through a thin synthetic silk fabric, spread over the surface of a solid medium. They could thus be separated from the substrate hyphae, which were collected after liquefaction of the medium. The amino acid and sugar content of hydrolysates of mycelia and cell walls from aerial and substrate hyphae was analyzed fluorimetrically by TLC. The characteristic of Actinomycetales, among the bacteria, is the tendency to branch. Mycelia are formed, which, within some families, e.g. Streptomycetaceae, differentiate into morphologically distinct aerial and substrate ones when cultivated on solid media. Biochemical differences between both types of mycelia are also to be expected: the substrate mycelium, above all, exchanges substances with the medium while the aerial mycelium forms spores. Distinct biochemical analyses however are rare, not least of all because of difficulties in separating aerial and substrate mycelia. In our work we tried to solve this problem. Materials and Methods Petri dishes (14 cm in diameter) were filled with Lab- Lemco-agar (Oxoid) containing 2 g/l yeast extract. The streptomycete strain to be analyzed was streaked on the surface of the medium in tight loops. The inoculum spores were harvested from the surface of well spored colonies. To obtain 100 mg of freeze dried aerial mycelium about 30 plates had to be prepared. To obtain 40 mg of cell wall material, which is just enough for the analysis of amino acids and sugars by the methods described below, more than 50 plates had to be incubated (Fig. 1). After inoculation the surface of the medium was covered with a piece of commercial synthetic silk. The silk fabric, which must be very fine, well stretched and uncoloured, was somewhat larger then the diameter of the petri dish and had been autoclaved beforehand in a separate dish. The silk got wet during the procedure but this was even advantageous for further handling. The fabric was then pressed down well on the surface of the medium using a sterile Drigalski spatula so as to eliminate any air bubbles. It must make contact with the medium right up to the edge of the petri dish so as to avoid the development of unseparable mycelia. In the course of incubation (28 C)the substrate mycelium developed below the silk fabric, while the aerial mycelium rose above it. For the separate harvesting of the two mycelial types first of all the aerial mycelium was scraped from the surface of the silk with a scalpel, washed and centrifuged repeatedly. During this treatment hyphal parts with no or few spores sedimented at 6,000 r.p.m.,while the hydrophobic spores floated on the surface of the washing water and could be decanted. The sedimented hyphal parts were then freeze- dried.
To collect the substrate hyphae the silk fabric was then removed and the petri dishes heated in steam until the agar dissolved. The nutrient agar was then decanted into heated centrifuge vessels and centrifuged for 5 min at 6,000 r.p.m. After pouring off the dissolved nutrient agar the sediment, containing the substrate mycelium, was washed with hot water at 80 C, heated for 30 min in steam and centrifuged again. This procedure was repeated twice to remove all the nutrient agar from the mycelium. The cleaned substrate mycelium was then freeze-dried in the same manner as the aerial mycelium and stored for biochemical analysis.
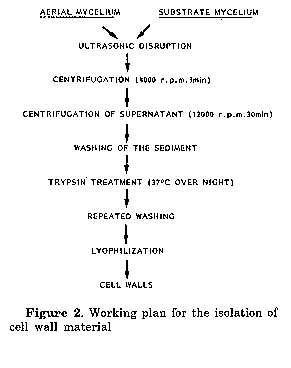
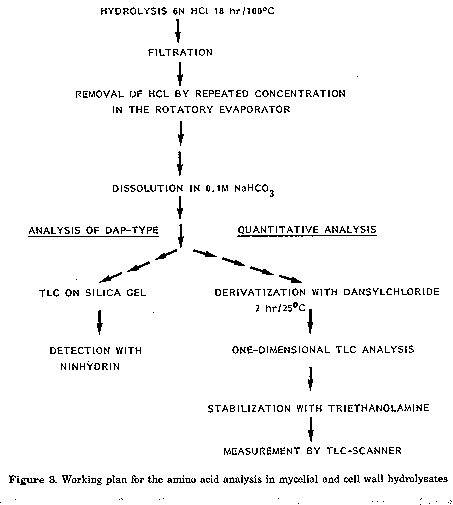
For the analyses whole cells and cell wall hydrolysates were used. For whole cell hydrolysates the freeze-dried hyphal material was used, cell wall fragments were prepared following the procedure described by Becker et al. (1965) and Yamaguchi (1965), except that the extraction step with ethanolic KOH was omitted, as well as the treatment with pepsin (Fig.2). The hydrolysis of whole cells and cell walls for amino acid analysis was carried out according to the usual methods (Cummins and Harris, 1956; Becker et al., 1965; Yamaguchi, 1965) disaggregating 20 mg of freeze-dried material with 1 ml 6N HCl for 18 hrs at 100 C in sealed lyophilization tubes. The hydrolysed material was then filtered, HCl evaporated from the tiltrate and the residue redissolved in 1 ml 0.1M NaHCO3 (Fig. 3). The amino acids were then derivatised with 5- dimethylamino-1-naphthalin-sulphonylchloride (dansylchloride) to give a yellow fluorescence. The treatment used was a combination of methods described by Gros and Labouesse (1969), Zanetta et al. (1970), Gray (1972) and Seiler and Wiechmann (1964). 0.5 ml of the hydrolysate in 0.1M NaHCO3 and 0.5 ml dansylchloride in acetone (10 mg/ml) were mixed in a tube which was then closed with a silicon stopper and allowed to stand for 2 hr in the dark until the solution was clear. Using a special apparatus the acetone was next evaporated via a tubule with nitrogen, while the residual dansylchloride settled and was removed by centrifugation. In both cases 2 ul of the supernatant were then spotted on two TL plates (Silicagel 60, Merck, 20x20 cm, 0.25 mm) with methyl acetate: isopropanol: NH4OH (45:35:20, v/v) and chloroform: methanoi: glac. acetic acid (75:20:5, v/v) and separated one dimensionally. After separation plates were dried and sprayed with a mixture of triethanolamine: isopropanol (1:4, v/v), dried in a dessicator for 16 hr and equilibrated at room temperature for 30 min before measurement. The quantitative analysis was carried out fluorimetrically in a TLC scanner (filters: primary 350-370 nm, secondary: 405 nm). The test for DAP-type was carried out according to Kroppenstedt (1979). For the quantitative analysis of sugars the hydrolysis of cell walls and whole cells was done following the procedure described by Becker et al. (1965) and Yamaguchi (1965) (Fig.4).
Chromatography of the monosaccharides was carried out on cellulose TLC sheets according to the method suggested by Kroppenstedt (1979), modified by 0.2M sodium acetate treatment to improve their separation. Development was carried out three times one dimensionally with ethyl acetate: pyridin: water (100:35:25, v/v). For the quantitative measurement of sugars the fluorescence method described by Klaus (1979) for the determination of sugars in wine and juice was used. After chromatography the dried sheets were dipped for 8-10 sec in a freshly prepared solution of 5 ml saturated Pb(IV)acetate and 5 ml of 1% 2,7-dichlorfluorescein in ethanol brought up to a volume of 200 ml with toluene. Monosaccharides as well as glycerol then showed a yellow-green fluorescence and were quantified with a TLC scanner under the same conditions as for the amino acids. Results and Discussion The method described gqves not only satisfactory results in separating aerial and substrate mycelia of streptomycetes, but also good quantitative values in spite of small amounts of material, because of the high sensitivity of fluorescence measurements (Neumeier, 1984).
One-dimensional chromatography in two solvents does not completely separate all the amino acids found in the mycelia, but allow them to be easily quantified in situ with a TLC scanner. This measurement gives a lower error than elution from TLC plates followed by measurement in cuvettes (Zanetta et al., 1970). The fluorescence intensity of the amino acid derivatives on the other hand fades with time, when it has not been stabilized with the procedure suggested by Seiler and Wiechmann (1966). The limit of detectability is 10^12 mol per component. For the separation of monosaccharides a modified gas chromatographic method (Pertzsch et al., 1981) was also used, which is well adapted to bacterial lysates but is time consuming, when a large number of cultures is to be tested, because of the purification by ion exchange chromatography. A further problem is selection of suitable nutrient media. They influence not only the biochemical composition and metabolism of an organism, but also the whole phenotype (formation of aerial mycelium, spores, colour, etc.). We used Lab-Lemco-agar with 2 g/l yeast extract. Its high protein content partly led to an atypical colony growth but it had two advantages: mycelia grew quickly and compact and sporulation was rare. In this way less sporulated aerial mycelia could be harvested and the spores could be separated by centrifugation, because of their hydrophobic character. Some other methods were tested for the separation of aerial and substrate hyphae: the plates were frozen after incubation and the aerial mycelium harvested with a scalpel or the silk was replaced by membrane filters and other foils, but none gave better results, though the silk is also very thick in relation to the height of the aerial mycelium so the lower parts remain embedded in the fabric. In these parts however the main differentiation and regulation processes that lead to the formation of aerial hyphae can be expected. Liquefaction of the solid medium by heat in order to harvest the substrate mycelium is rather crude and cannot be applied for physiological studies, e.g. enzymatic activities. We tested gelatine instead of agar but all the strains investigated liquefied it during incubation. On the other hand agarase is too expensive. References Becker, B., M.P. Lechevalier & H.A. Lechevalier (1965). Chemical composition of cell wall preparations from strains of various genera of aerobic actinomycetes. Appl. Microbiol., 13:236-243 Cummins, C.S. & H.Harris (1956). The chemical composition of the cell wall in some gram-positive bacteria and its possible value as a taxonomic character. J. gen. Microbiol., 15: 583-600 Gray, W.R. (1972). End-group analysis using dansylchloride. Methods in Enzymology, 25: 121-138 Gros, B. & B.Labouesse (1969). Study on the dansylation reaction of amino acids, peptides and proteins. Eur. J. Biochem., 7:463-470 Klaus, R. (1979). Quantitative DC-Analyse von Zuckern, Zuckersauren und Polyalkoholen. Kontakte, 3/79:24-27 Kroppenstedt, R.M. (1979). Chromatographische Identifizierung, dargestellt am Beispiel der Actinomyceten. Kontakte, 2/79:12-16 Neumeier, W (1984). Aminosauren- und Zuckergehalte in verschiedenen Mycelformen bei Streptomyceten. Dissertation, Giessen, BRD Pertzsch, C., L. Bender & K.H. Neuraann (1981). Radiogaschromatographie von Kohlenhydraten, organischen Sauren und Aminosauren aus Pflanzen. Z.Pflanz. Bodenkde., 144:231-240 Seiler, N. & J.Wiechmann (1964). Zum Nachweis von Aminosauren im 10-10 Mol-Masstab. Trennung von 1-Dimethylaminonaphthalin - 5-sulfonyl-aminosauren auf Dunnschichtchromatogrammen. Experientia, 20:559-560 Seiler, N. & J.Wiechmann (1966). Quantitative Bestimmung von Aminen und Aminosauren als 1-Dimethylamino-naphthalin-5- sulfonsaureamide auf Dunnschichtchromatogrammen. Z.Anal. Chem., 220:109 Yamaguchi, T. (1965). Comparison of the cell wall composition of morphologically distinct actinomycetes. J. Bacteriol., 89:444453 Zanetta, J.P., G.Vincendon, P.Mandel & G.Gombos (1970). The utilization of 1-dimethylaminophthalene-5-sulphonyl chloride for quantitative determination of free amino acids and partial analysis of primary structure of proteins. J. Chromatogr., 51: 441-458. Copyright 1990 C.E.T.A., The International Centre for Theoretical and Applied Ecology, Gorizia
The following images related to this document are available:Line drawing images[ac90003b.gif] [ac90003d.gif] [ac90003c.gif] [ac90003a.gif] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}