 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Associations Of Actinomycetes With Algae L.V.KALAKOUTSKII *, G.M.ZENOVA, V.S.SOINA and A.A.LIKHACHEVA *Institute of Biochemistry and Physiology of Microorganisms, USSR Academy of Sciences, Pushchino and Chair of Soil Biology, Dept. Soil Science, Moscow State University, Moscow, USSR
Code Number: AC90006
Sizes of Files:
Text: 61.8K
Graphics: Photograph (jpg) - 155.9K ABSTRACT. The review focuses on the behaviour of streptomycetes and a green eukaryotic alga (Chlorella) in mixed cultures. Possibility of establishing common lichen-like thalli is confirmed; favourable cultural conditions are specified and distinctive properties of resulting associations documented. Compatible partners could be chosen by simple tests, developed to detect positive tropism of actinomycetes towards algae, as well as to monitor changes in cosedimentation patterns in mixtures of actinomycete and algal cells. Pairs displaying reciprocal or one-sided antagonism were noncompatible; growth stimulation was noted with effective combinations of partners. Natural communities of algae and bacteria, with predominance of streptomycetes, were found of some outcrops on carbonaceous rocks in temperate climates. A bibliography on behaviour of mixed cultures of actinomycetes with other organisms is included. Like other bacteria, actinomycetes are able to form associations with various organisms under natural and/or laboratory conditions. Such associations differ in intensity and nature depending on the mutual effects of the partners, as do their stability and significance and consequently the coverage they received in the literature. Due to their applied significance, associations of pathogenic actinomycetes with animals (e.g., see Beaman, 1981) and of actinomycetes of the genus Frankia with higher plants yielding nitrogen-fixing symbioses (e.g., see Becking, 1961; Benson, 1988; Berry, 1986) are better studied. Streptomycetes and nocardiae causing plant diseases as well as symbionts of insects (Goodfellow and Williams, 1983) are less studied. In the past knowledge of physiological and morphological properties of the actinomycetes was derived mainly from studies of pure laboratory cultures. This is especially true of representatives of the genus Streptomyces intensively studied during the era of antibiotics. It was the insistence on studies with pure cultures which probably led Waksman (1959) to question their relevance in the interpretation of the activities of actinomycetes in nature. Even a cursory inspection of information summarized in Table 1 definitely shows that under conditions of associative growth, not unexpectedly, the performance of partners is often modified. Frequently the actually observed changes cannot be predicted from investigations on pure cultures. Studies on the establishment and behaviour of associations may thus reveal properties of actinomycetes which for various reasons had escaped attention. While attempting to experimentally construct and study associations of streptomycetes with protists our attention was attracted to the possibility (Lazo, 1964, 1966) of establishing an association of actinomycetes and algae so as to develop an "actinolichen" under laboratory conditions. Reasons for this interest may be summarized as follows. - Associations of this type are little studied; there is almost no information on their properties or whether they exist in nature; however there are no reasons to deny a priori the possibility of their occurrence. Even the experimental construction of such associations might open up an unusual perspective in the field of lichenology and studies on the evolution of symbioses. - Eukaryotic algae are plants, though simple and their interrelationships with the actinomycetes are currently being closely studied (see reviews cited above). Thus the development of simple criteria for testing compatibility of plantactinomycete associations seemed relevant. - Studies on "classical" natural lichens (e.g., associations of a mycobiont and an algobiont) often demonstrate that peculiar metabolic and bioactive products can be synthesized (e.g., antibiotics of the group of lichen acids, Cluberson et al., 1984; Peveling, 1987) which had not been detected when the partners were cultivated separately. Table 1. Reported examples of associations of actinomycetes with other organisms. Although preliminary selection of partners is usually mentioned, tl experimental procedures actually used for selection are only occasionally specified.
Partner 1 Partner 2 Association formation
------------------------------------------------------------
Streptomyces Bacterium Preliminary selection
griseus coli, Bacillus of partners
subtilis, S.
longisporus
S. albus Aspergillus Mixed culture
niger,
Fusarium
culmorum
S.scabies Ceratostomella Preliminary selection
ulmi, of partners
Verticillium
dahliae,
Helminthosporium
victoriae,
F. oxysporum
f. lycopersici
Streptomyces sp. Corynebacterium equi Inoculation of bacteria
into growing
actinomycetes
S. griseus Penicillium sp. Preliminary selection
of partners
S. griseus B. mycoides Preliminary selection
of partners
S. aureus Gram-positive Preliminary selection
bacteria of partners
S. aureus S.griseus Preliminary selection
of partners
S. lavendulae Streptomyces sp. Preliminary selection
of partners
S. verne Streptomyces sp. Preliminary selection
of partners
S. griseus S.verne Preliminary selection
of partners
S. aureofaciens Torulopsis sp. Preliminary selection
of partners
Micromonospora Bacillus sp. A laboratory contaminant
S.coelicolor B.rusticus,Bact. Filtrates of cultural
liquefaciens,Bact. broth of bacteria added
nigrificans, to actinomycete culture
Achromobacter
agile
Streptomyces sp. Chlamydomonas sp. Experimental biotic
comunity found by
screening of two
membered combinations in
soil-water cultures
Strepto-
verticilluim sp. Streptomyces sp. Preliminary selection
of partners
S. malachito-
spinus Phycomyces Preliminary selection
blakesleeanus of partners
S.levoris,
S. nodosus Candida tropicalis, Preliminary selection
C. utilis, C. of partners
guilliermondii,
C. krusei,
Saccharormyces
cerevisiae
Stv.mycohepti-
cinicum Schizosaccharomyces Preliminary selection
rolnys, Rhodotorula of partners
mucilaginosa, C.
utilis
S. sindenensis Rhodococcus sp. A stable laboratory
association
Coryneform
bacterium Chlamydomonas Bacterium isolated as
rheinardtii contaminant of a
laboratory algal culture
Nocardia minima Arthrobacter citreus Mixed culture in
artificial ecosystem
S. purpurtus Bacillus sp. Mixed culture in
artificial ecosystem
S.levoris C. tropicalis Preliminary selection
of partners
S.rimosus S.violaceus, S. Preliminary selection
violocinereus of partners
Table 1 continued
Partner 1 Partner 2 Some phenomena thought
to be characteristic
of the association
--------------------------------------------------------------
Streptomyces Bacterium Intensification of anti-
griseus coli, Bacillus biotic biosynthesis
subtilis, S.
longisporus
S. albus Aspergillus Changes in sensitivity
niger, to antibiotics in fungi
Fusarium
culmorum
S.scabies Ceratostomella Intensification of
ulmi, pigment production in
Verticillium fungi
dahliae,
Helminthosporium
victoriae,
F. oxysporum
f. lycopersici
Streptomyces sp. Corynebacterium equi Intensification of
antibiotic biosynthesis
S. griseus Penicillium sp. Intensification of
antibiotic biosynthesis
and modification of
cultural properties of
actinomycetes
S. griseus B. mycoides Change in pigmentation
of actinomycete
colonies,
intensification of
antagonism
S. aureus Gram-positive Restoration of the
bacteria ability of
the actinomycete to
synthesize the
antibiotic antimycoin
S. aureus S.griseus Restoration of ability
to form spores
S. lavendulae Streptomyces sp. Restoration of ability
to form spores
S. verne Streptomyces sp. Restoration of ability
to form spores
S. griseus S.verne Restoration of ability
to form spores
S. aureofaciens Torulopsis sp. Restoration of ability
to form spores
Micromonospora Bacillus sp. Enhanced actinomycete
growth and
intensification of its
antibiotic activity
S.coelicolor B.rusticus,Bact. Intensification of
liquefaciens,Bact. antagonistic properties
nigrificans,
Achromobacter
agile
Streptomyces sp. Chlamydomonas sp. Mutualistic stimulation;
akinetokinesis
induction, increades
motility and decreased
extracellular
polysaccharide
production in the alga;
enhanced sporulation in
the actinomycete
Strepto-
verticilluim sp. Streptomyces sp. Stimulation of pigment
production of
streptomycete
S. malachito-
spinus Phycomyces The actinomycete
blakesleeanus produces an active
substance presumably
taking part in the
differentiation of
intracellular
structures at the time
of gametangium formation
S.levoris,
S. nodosus Candida tropicalis, Intensification of bio-
C. utilis, C. synthesis of antibiotics
guilliermondii, (levorin and
C. krusei, amphotericin B).
Saccharomyces Inhibition of yeast
cerevisiae growth. Synthesis of
lytic enzymes by the
actinomycetes enhanced
Stv.mycohept-
icinicum Schizosaccharomyces Intensification of bio-
rolnys, Rhodotorula synthesis of mycoheptin,
mucilaginosa, C. inhibition of yeast
utilis growth
S. sindenensis Rhodococcus sp. Formation of "fruiting
bodies", presumably as a
result of morphological
alteration of the
organism in the
association
Coryneform
bacterium Chlamydomonas Preferential synthesis
rheinardtii by the bacterium of a
single protein (m.w.
21,500) following
exposure to light
Nocardia minima Arthrobacter citreus Intensification of
synthesis of trombolytic
proteases and increased
polymorphism in N.minima
S.purpuratus Bacillus sp. Intensification of
synthesis of
neopurpuratin
S.levoris C. tropicalis Intensification of
synthesis of levorin,
levoristatin and of
fatty acids
S.rimosus S.violaceus, S. Increased yield of
violocinereus proteases with fibrino-
and trombolytic
activity. In S. rimosus
growth processes
disturbed (intra-hyphal
hyphae), in S.
violocinereus
hypersynthesis of
peptidoglycan, cell wall
thickening, mesosome
hypertrophy, loss of
ability to form
capsules, spore
formation retarded.
Table 1 continued
Partner 1 Partner 2 References
------------------------------------------------------------
Streptomyces Bacterium Krasilnik et al., 1951
griseus coli, Bacillus
subtilis, S.
longisporus
S. albus Aspergillus Rehm, 1953,1959
niger,
Fusarium
culmorum
S.scabies Ceratostomella Grossbard 1955
ulmi,
Verticillium
dahliae,
Helminthosporium
victoriae,
F. oxysporum
f. lycopersici
Streptomyces sp. Corynebacterium equi Craveri,1956
S. griseus Penicillium sp. Aleshina and
Makarovskaya.,1956
S. griseus B. mycoides " " "
S. aureus Gram-positive Bush, 1956
bacteria
S. aureus S.griseus Dondero & Scotti, 1957
S. lavendulae Streptomyces sp. "" ""
S. verne Streptomyces sp. "" ""
S. griseus S.verne "" ""
S. aureofaciens Torulopsis sp. Belik et al., 1957
Micromonospora Bacillus sp. Egorov et al., 1960
S.coelicolor B.rusticus,Bact. Egorov et al., 1960
liquefaciens,Bact.
nigrificans,
Achromobacter
agile
Streptomyces sp. Chlamydomonas sp. Parker & Bold, 1961
Strepto-
verticillium sp. Streptomyces sp. Tresner et al. 1971
S. malachi-
tospinus Phycomyces Kozlova et al. 1971;
blakesleeanus Preobrazhenskaya
et al., 1975
S.levoris,
S. nodosus Candida tropicalis, Yakoleva et al 1972,
C. utilis, C. 1981,1985;Yakoleva,1983
guilliermondii,
C. krusei,
Saccharormyces
cerevisiae
Stv.mycohept-
icinicum Schizosaccharomyces Kuznetsova, 1974
rolnys, Rhodotorula
mucilaginosa, C.
utilis
S. sindenensis Rhodococcus sp. Locci & Locci, 1975
Coryneform
bacterium Chlamydomonas Stegeman & Hoober, 1975
rheinardtii
Nocardia minima Arthrobacter citreus Egorov &Landau, 1982
S.purpuratus Bacillus sp. Ohshima et al., 1983
S.levoris C. tropicalis Kuznetsova et al., 1984
S.rimosus S.violaceus, S. Egorov & Landau, 1986
violocinereus.
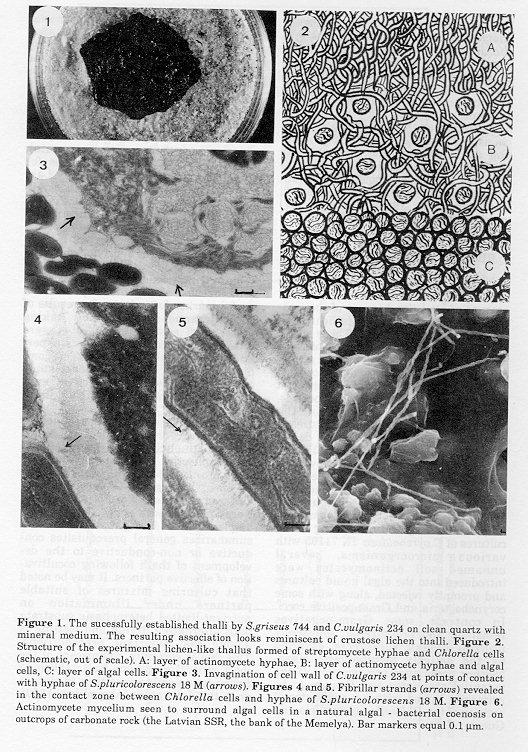
Results And Discussion Among the green algae, Chlorella spp. probably received maximum amount of attention because of their potential application and significance in contamination problems. These algae are also prominent on account of their ability to enter many and diverse symbiotic associations (Smith and Douglas, 1987). However the extensive number of bacterial contaminants and associates, found by various workers in the laboratory as well as in mass cultures of diverse Chlorella strains, usually do not mention representatives of the Actinomycetales (for a review, see Gitelson et al., 1981); neither are these recorded among the likely bacterial symbionts of Chlorella (Nakamura, 1963). In a study, aimed at experimentally establishing and monitoring mixed cultures of C. pyrenoidosa TX 71105 with various microorganisms, several unnamed soil actinomycetes were introduced into the algal liquid cultures and promptly rejected, along with some corynebacteria and Gram-positive cocci. In contrast, some Gram-negatives remained and proliferated, selectively utilizing organic substances, excreted by the actively metabolizing algae. Neither acceptance nor rejection of the potential associates was thought to be primarily linked with an antibacterial activity of the Chlorella strain (Vela and Guerra, 1966). Although a mention of an association between actinomycetes and a soil alga, Chlamydomonas sp., involving likely cooperation probably dates back to the paper by Parker and Bold (1961), it was the work of Lazo (1964, 1966) which provided the first evidence that the experimental construction of an association resembling lichen, but involving an eukaryotic alga and prokaryotic mycelial organism (streptomycete), might indeed have some chances for success. By using light microscopy Lazo (1964, 1966) demonstrated the establishment of a lichen-like thallus ("actinolichen") in mixed cultures of C. xanthella and streptomycetes of the Cinereus series ("species group"). It seems, that the morphological distinctiveness of the thallus formed under certain conditions and with certain combinations of organisms provides a useful marker for judging the results of the interactions, presumably taking place when some actinomycetes were mixed with algae. This criterion permits the visual assignment of combinations of the organisms under study to either "effective" or "ineffective", even in the absence of other possible criteria of compatibility. In an extension of studies aimed at establishing, under laboratory conditions, actinolichen-type associations, several effective combinations, involving C. vulgaris 234 and Streptomyces species, belonging to the sections Cinereus, Helvolo-Flavus and Albus, were found (Zenova et al., 1980). An example is shown in Fig. 1, while Table 2 summarizes general prerequisites conductive or non-conductive to the development of thalli following cocultivation of effective partners. It may be noted that culturing mixtures of suitable partners under illumination on nutrient-limited heterophasic substrata with alternating drying and wetting is among the important preconditions. This fact is certainly reminiscent of the most general conditions for establishing lichen-type symbioses either in nature or in the laboratory (Ahmadijan, 1980).
Table 2. Conditions for the establishment of the experimental lichen-like thallus.
CONDITIONS UNDER WHICH THE CONDITIONS UNDER WHICH THE
THALLUS IS ESTABLISHED THALLUS IS NOT ESTABLISHED
SUBSTRATUM
Clean quartz sand wetted Liquid and solid media
with suitable mineral traditionally used for
medium, potato broth or laboratory cultivation
sterile tap water; potato of either algae or
blocks immersed in water streptomycetes
INOCULUM
Algal cells (7-10 days) Considerable predominance
grown with illumination of streptomycete mycelium in
in mineral medium, the inoculum, but not vice versa
separated from the medium;
Streptomycetes mycelium
(3-5 days) grown in a
submerged culture in potato
broth, washed off from the
medium (Recommanded ratio
"algal cells: streptomycete
mycelium" 1:1)
CULTURE CONDITIONS
Luminostat, intensity of Thermostat, incubation in
continuous illumination: darkness. Excessive wetting
8-10,000 lux, temperature: or drying of the substratum
28-30 C. Initial wetting under conditions of either
of the substrate up to 60 % illumination or darkness.
of the total moisture
capacity, followed by slow
drying and rewetting
after 3-5 days.
In a sense the inductive conditions appear not to be optimal for vegetative growth of either the alga or the actinomycete able to form a thallus. Alternate drying and wetting appears to be critical. Using light microscopy it was possible to reveal the laminated structure of the experimental lichen-like thallus (Fig. 3). The following layers were clearly distinguishable: the actinomycete hyphae layer, that of intertwining actinomycete hyphae and algal cells and the layer of algal cells. Electron microscope investigations of ultrathin sections of the experimental thalli demonstrated that within the thallus the algal cells, as a rule, undergo a normal cycle of division, growth and dying off. In streptomycetes no spore formation was noted microscopically; absence of spores was confirmed by the lack of growth on oatmeal agar inoculated with a thallus suspension preheated at 55 C for 10 minutes. In the zone of mixed growth a close contact between hyphae and algal cells was apparent. In algal cells contacting actinomycete hyphae some modifications were noted: invagination of the Chlorella cell wall at the point of contact, contraction of the protoplast, occurrence of a rather wide periplasmic space between the plasmalemma and the cell wall of the alga (Fig. 4). Fibrillar strands were noted where contact occurred between streptomycete hyphae and algal cells (Fig. 5). The overall picture of the contact zone was highly reminiscent of that observed in several natural lichens (Cock, 1977). On rather rare occasions streptomycete hyphae were seen to penetrate the algal cell wall and closely adjoin the cytoplasmic membrane, the latter undergoing invagination. In these cases modifications in the contacting cells may be considerable and include partial destruction of the algal cell wall; simultaneous occurrence of a double cell wall in the contacting streptomycete hypha, possibly as result of intrahyphal hyphae formation or operation of some other mechanism, was noted. In the experimentally established thallus dead algal cells filled with streptomycete hyphae were very rarely seen. The results of light as well as electron microscopy thus suggest that within successfully established and morphologically diversified associations the prevailing relationships of constituent partners look quite balanced. They hardly exemplify a mere coexistence in a common space. Frequent intimate contacts between actinomycete and algal cells suggest the possibility of some metabolic interrelationships worthy of further consideration. Failure to sporulate in all streptomycetes so far studied within established thalli suggest that the cooperation in question might involve not only possible "gains" but also concominant, albeit temporary, "losses". Experimentally constructed thalli showed a clear ability to reproduce as distinctive entities under proper conditions. Portions of the thallus, formed on the surface of quartz sand and transferred into a suitable fresh substratum (a potato block immersed in water or the surface of wetted quartz sand), developed into a new thallus similar to the initial one. A lichen-like thallus persisted for five or more transfers. To separately assess the actinomycete and algal biomass within thalli, total amounts of diaminopimelic acid (DPA) and chlorophyll were determined (Robinson, 1968) and converted into respective cell biomasses using data of parallel determinations with pure cultures. Although the resulting figures might not be very precise, they nevertheless appeared useful in various contexts. The relative contribution of actinomycete biomass in a number of constructed thalli appeared to vary within the range of 60-90%. This is rather similar to the range of figures given in the literature (see, e.g., Millbank, 1976) for the relative contribution of the mycobiont biomass in natural lichens. On the other hand the proportion of actinomycete biomass in "mature" thalli differed from the one found to be optimal (Table 2) at the mixing and initiation stage. There is some evidence that vegetative cells in the experimental thalli are better protected against the damaging effect of dessication, as compared to separate cultures (Zenova et al., 1986). Thus exposure of pure cultures of the streptomycete, of the algae and of the lichen-like thallus formed by S. griseus 744 and C. vulgaris 234 to conditions of low humidity (aw = 0.32) for 40 days (24 C, in a luminostat) demonstrated that in the pure cultures the survival rate of vegetative cells of the streptomycete (counted as colony forming units following standard disaggregation treatments) was 16 % and 6% for the alga, while in the thallus the values were 59% and 7% respectively. The thalli were also superior in terms of dessications survivability to mechanical mixtures of corresponding streptomycetes and algal cells. Once the thalli had been successfully established, it was usually possible to maintain them in an air-dry state for more than 12 months. Photosynthesis seemed to resume in dried thalli on rewetting at aw = 0.96. Several examples were found demonstrating that the apparent antimicrobial activity of the actinolichen-type associations might differ from that of its individual components cultured separately under various sets of experimental conditions (Zenova et al., 1983). An interesting tendency observed was the enhancement of antagonistic abilities against Gram-positive bacteria in general and various representatives of Actinomycetales in particular. Thus a thallus formed by C. vulgaris 234 and S. griseus 744 (both strains are antagonistically inactive in pure culture) showed antagonistic properties against Staphylococcs aureus, Arthrobacter globiformis, S. fradiae and S. antibioticus. There are possibly some substances among the metabolites of C. vulgaris 234 excreted into the cultural liquid even under conditions of separate cultivation, which exert a favourable effect on the apparent antagonistic potential of some streptomycetes. Thus when a number of streptomycete strains were cultured on water agar media supplemented with lyophilized Chlorella broth supernatant, antagonistic abilities were demostrated by the agar-block method in S.griseolus 140, S. carpaticus 463, S. bikiniensis 2499 against several representatives of Actinomycetales (excluding mycobacteria) and streptomycetes of the Cinereus seties. Less or no activity was revealed against the same test organisms when the above-mentioned streptomycetes were cultured either on Gromov's (1965) mineral medium supplemented with individual polyols (0.1% w/v of dulcitol, sorbitol, inositol, erythritol, mannitol, ribitol, arabitol) or on complex organic media (peptone-corn agar). To make comparisons possible, in the experiments of these series the apparent antibiotic activity measured by the agar diffusion method was related to the biomass unit of the antagonistic streptomycetes as represented by total protein content per standard agar block. Whether the effect observed had anything to do with the tendency of excluding some "third partners" from the developing thalli, or what might be the nature of the substance(s), excreted by Chlorella which apparently favours the expression of antagonistic activity by streptomycetes, remains to be seen. Among other features expressed by streptomycetes in the established associations with the alga one might mention the enhancement of apparent extracellular enzyme activities, clearly demonstrable on occasions where no similar activities were expressed in pure cultures under diverse experimental conditions. Such was the case, for instance, with the activity of extracellular amylases recorded in associations involving S. griseolus 140 and S. bikiniensis 2429. The first attempts to construct in vitro lichen- like associations of actinomycetes with algae were based mainly on a "trial-and-error" approach. Results of successful trials revealed the generally "inductive" conditions of co- cultivation as well as providing important hints on the optimal proportions of partners cells in successful mixtures. In addition prescreening of prospective partners for reciprocal antagonistic activity serves a useful purpose. Pairs in which at least one partner showed antagonistic activity toward the other (agar block method) were inefficient at establishing thalli. Since a separate estimation of algal and streptomycete biomass in the established thalli was possible using chlorophyll and DPA as markers, the respective biomass yields were compared in 7 day old thalli and in pure cultures of the algae and streptomycetes potentially able to enter associations. Both were grown on clean quartz sand wetted with Tamyia's (1953) medium for the same period with or without illumination. In the case of effective partners the respective biomass yields in the successfully established thalli tend always to be higher than in separate cultures. For instance the biomass of S. griseus 744 in the successfully established thalli was 51 mg, against 32 mg in separate culture. The total biomass yields of C. vulgaris 234 in thallus was 1.2 mg vs. 0.8 mg in pure culture. To obtain these results equal inoculum of algal cells and streptomycete mycelium for thalli and pure cultures were compared. The reasons seem to be obvious for the heterotrophic partner, presumably feeding on algae excretions and lysis products. The reasons for the apparent stimulation of algal growth, observed in several experiments are not so clear. Immediate suggestions would stress a possible increase in CO2 concentration in the vicinity of algal cells and/or removal of growth-inhibiting metabolites. There is of course a possibility that the relative content of either chlorophyll or DPA per biomass unit might vary with the physiological conditions prevailing and hence the accurate comparative estimations of biomass yields would be possible only using additional independent criteria. Yet judging from the "healthy visual appearance" of the successfully developing thalli, the possibility of algal stimulation through some sort of metabolic cooperation should probably not be excluded a priori. All the above data notwithstanding, the fact remains that in the course of a "trial-and-error" approach to the construction of lichen-like thalli one was always confronted with combinations (pairs) of organisms, some of which appeared to be successful in establishing the thalli under appropriate conditions, while others were not. Among the tests mentioned above only the probe for reciprocal antagonism proved useful in excluding from further consideration apparently hopeless combinations. Yet inefficient combinations included also some pairs with no apparent symptoms of mutually antagonistic activities. Therefore some simple techniques were sought which might be helpful in preliminary screening aimed at finding combinations of compatible organisms. It was hoped that if such techniques would really work in predicting effective partners, even the interim results of their application might shed additional light on diverse aspects of processes likely to be involved in the establishment of specific reletionships between compatible partners. One of the techniques employed is the membrane filter penetration test. The experiments took advantage of the known ability of streptomycete hyphae to penetrate through the pores in membrane filters (Polsinelli and Mazza, 1984), while growing against the gradient of nutrient concentration. In the actual experiments the streptomycetes tested were inoculated by means of a sterile spatula into the surface of tap water agar. Algal cell slurry samples were placed on sterile membrane filters (pore size d=1.5 um). Filters bearing algal cells along with sterile controls were placed on actinomycete lawns (algal layer up), the Petri dishes were closed and incubated under illumination or darkness at 28 C for 7 days. Following incubation the filters were removed, washed, fixed, stained with 0.3% (w/v) erythrosine solution and examined under the oil immersion system. Relative abundance of streptomycete hyphae (total hyphal length) appearing on the unit area of the filter surfaces directly in contact with algal cells was estimated and compared to the control. The results appeared to be reproducible. Several streptomycetes showed positive tropism towards selected algal strains, either in the light or in darkness, others under both conditions and finally some were almost completely indifferent. A possible inference was that streptomycetes showing positive tropism under illumination tend to feed on product(s) excreted by actively metabolizing algae, whereas those showing positive tropism in darkness utilize product(s) leaking from algal cells deprived of an energy source. Positive tropism of some kind was invariably found with streptomycetes shown in separate tests to be able to participate in the establishment of lichen-like thalli. Interactions involving recognition of suitable partners in microorganisms often occur (e.g., Mirelman, 1986) through enhancement of aggregation followed by cosedimentation from mixed suspensions. An attempt was made to find whether interactions of this sort do indeed take place on mixing suspensions of certain algal and streptomycete cells (Likhacheva et al., 1987). For the experiments algae were grown in Tamyia's medium for 7 days and resuspended in the same medium or in phosphate buffers (pH 7.0-9.0). Streptomycetes were grown on potato broth in submerged cultures and harvested at the beginning of the stationary growth phase. To quantify results of experiments, suitable suspensions of untreated or treated streptomycete and algal (10^6 cells/ml) cells were mixed and allowed to sediment for fixed time intervals (40 min) in 5 mm wide test tubes. Test tubes with the resulting sediments were projected onto a screen and sediments measured. Comparisons of suspension behaviour were made using a simple aggregation index:
K=hm.ma/ha.mm
where hm and mm stand for sediment height (h) and dry weight
(m) in mixtures, ha and ma for sediment height and dry weight
respectively in pure actinomycetes cultures.As visual and light microscopic observations suggested, there were appreciable differences in the sedimentation behaviour of various pure and mixed suspensions of streptomycetes and algal cells. In some cases the rate of sedimentation was enhanced. The sediments in some mixed suspensions appeared to be double-layered, streptomycete hyphae being found mainly at the bottom, while algal cells formed the top layer. In other combinations the settled material was composed of aggregates representing rather homogeneous mixtures of streptomycete hyphae and algal cells. In the case of aggregate formation the maximal radii of settling particles calculated according to Stokes' equation (Vadjunina and Korchagina, 1986) for experimental aggregates in mixed suspensions exceeded those theoretically expected for simple mechanical mixtures of the streptomycete and algal cells. Experimental observations suggested that interactions possibly involving recognition and certainly involving alterations in cosedimentation patterns, as a result of aggregate formation, do indeed occur in certain mixtures of streptomycetes and algal cells. The important point, however, was a significant correlation between compatibility of partners, as judged by their ability to establish thalli, and occurrence of positive tropism on the one side, and sedimentation patterns in mixed suspensions, as represented by increased values of aggregation index on the other. In actual experience the aggregation indexes varied significantly, but with compatible partners they usually were >1.0, whereas with noncompatible <1.0. Since the experiments on cosedimentation were the easiest to perform, they appeared to be a simple but valuable tool in prescreening for compatible partners. In contrast to Gram-negative bacteria, only scant information is available on the likely mechanisms involved in the recognition of compatible partner(s) and attachment in actinomycetes. Mention has been made of the possible involvement of lectins in streptomycetes in connection with hemoagglutination (Fujita et al., 1972; Kamegama et al., 1982), in interactions of microaerophilic Actinomyces spp. with other bacteria inhabiting the oral cavity (Cisar, 1986), as well as in Frankia spp. capable of recognizing suitable plant host roots (Chaboud and Lalonde, 1983). As results of some of the observations showed (Table 3), the structures of the streptomycete cells responsive to interaction with algae in cosedimentation experiments seem to be very delicate indeed, responsivity being damaged even when centrifugation was employed in routine washing procedure. Table 3. Effect of washing protocol of actinomycete hyphae on the rate of their cosedimentation with C. vulgaris 234 cells. Experimental figures represent values of the aggregation index.
Streptomycete FILTRATION IN CENTRIFUGATION IN
species Phosphate Potato Phosphate Potato
and strains buffer(pH) broth buffer(pH) broth
--------------------------------------------------------------
4.5 7.0 9.0 4.5 7.0 9.0
--------------------------------------------------------------
S.griseolus 140 0.80 1.40 1.50 1.70 0.60 0.50 0.70 0.65
S. bikiniensis 2429 1.00 1.20 1.80 1.85 0.90 0.83 0.83 0.40
Exposure to an acidic environment (pH 4.5) was also detrimental. There is thus some likelihood that the responsive structures are associated with the surface of the streptomycete cells. The interactive ability of selected streptomycete cells with C. vulgaris 234 (Table 4) in cosedimentation experiments was clearly inhibited by prior heating (85 C for 30 min) and treatment with 1% w/v trypsin (1 h) which indicates a possible involvement of protein molecules. Among the sugars tested in pretreatment experiments on their ability to block aggregation, maltose appeared to be prominently effective. The results thus do not contradict the suggestion of streptomycete lectin(s) involment in interactions with algal cells, although by no means prove it. Table 4. Effect of heating and of pretreatment with either trypsin or maltose on aggregational behaviour of streptomycetes mixed with cells of C.vulgaris 234. Figures stand for aggregation indexes found experimentally. For untreated controls see Table 5.
Streptomycetes Heating Trypsin Maltose ------------------------------------------------ S. griseolus 140 0.74 0.85 0.57 S. pluricolorescens 18M 0.45 0.49 0.58 S. alboniger 462 0.41 0.50 0.53 S. carpaticus 463 0.48 0.50 0.61 S.bikiniensis 2429 1.00 0.76 0.90 S.globisporus 41 0.60 0.70 0.87 S.fimicarius 295 0.61 0.83 0.66 S.globisporus 92 0.80 1.00 0.80 Table 5 illustrates results of experimental screening for compatibility and ability to establish thalli in several streptomycetes and C. vulgaris 234. A note should be made both of the results as well as of the dependability (i.e., predictive value) of individual tests related to growth stimulation, positive tropism and cosedimentation patterns. As may be seen, streptomycetes isolated from various sources and found to be effective in the ability to establish lichen-like thalli with C. vulgaris belong mainly to the series (formerly thought to represent "species groups") Cinereus, Helvolo-Flavus and Albus; two streptomycetes belong to cluster IB, S. albidoflavus (Williams et al., 1983). The experimental construction of algal-actinomycete thalli under laboratory conditions does not prove, of course, the existence of similar associations under natural conditions. The results merely suggest the possibility that the interactive steps mentioned are also involved on occasions when algae and actinomycetes have the chance to coexist under natural conditions. It has been reasoned that such chances might present themselves in algal blooms, appearing on the outcrops of carbonaceous rocks (characteristic neutral reactions and the abundance of calcium presumably making coexistence of actinomycetes possible). Such blooms were shown indeed (Zenova et al., 1988) under conditions of a temperate climate to consist of algal - bacterial communities of a rather distinctive character. Using scanning electron microscopy of undisturbed samples, luminescent microscopy and platings on appropriate media with subsequent identification of the main communities components, several groups were revealed. Those with the following dominant algal groups were prominent: 1) Nostocales; 2) Nostocales + Chlorococcales; 3) Nostocales + Oscillatoriales; 4) Oscillatoriales + Chlorococcales. No fungal hyphae were observed in the intact communities. Among the heterotrophic organisms present vegetative hyphae of actinomycetes and certain Gram-positive bacteria (rhodococci and micrococci) predominated (especially in cases 1 and 4) if assayed in terms of either relative biomass contribution or total colony forming units. Table 5. Examples of effective vs. noneffective interactions of selected streptomycetes with C.vulgaris 234. L and D = streptomycete tropism towards the alga in the light and in the dark respectively. Assignement to section, series and species was made according to Gauze et al. (1963). Streptomycetes isolation source: cinnamonic carbonate soil (140, 2429, 295, 2C15), takyr-lake solonetz (18M, 41, 92, 19,429, K63), algologically pure culture (463) and crustose lichen (26, A234). (CA=Cinereus Achromogenes; H- FH=Helvo-Flavus Helvolus; CC=Cinereus Chromogenes; AA=Albus Albus; RF=Roseus Fuscus)
Section Species Stimultion Tropism Aggre- Thallus and and of growth of towards gation forma- series strain partners alga index tion -------------------------------------------------------------- CA S.griseolus 140 + L, D 1.75 + H-FH S.pluricolorescens18 + L, D 2.15 + CC S.carpaticus 463 + L, D 2.70 + CA S.bikiniensis 2429 + L, D 1.70 + H-FH S.globisporus 41 + L 1.05 + " S.fimicarius 295 + L 1.80 + " S.globisporus 92 + D 1.80 + AA S.alboniger 462 + D 2.21 + H-FH S.helvoloviolaceus19 - - 0.77 - RF S.moderatus 2015 - - 0.13 - CA S.aureofaciens 429 - - 0.56 - H-FH S.chrysomallus K63 - - 0.50 - CC S.xanthocidicus 26 - - 0.89 - " S.roseogriseus A234 - - 0.26 - No actinolichen-like associations similar to those constructed in the laboratory have been found so far. However, pictures of algal cells immersed in the network of closely contacting actinomycete hyphae (Fig. 6) were quite infrequent. It is perhaps noteworthy that among dominating streptomycete species, isolated from natural algo-bacterial communites 1-4, representatives of the following series were prominent: Albus, Helvolo-Flavus, Cinereus and Roseus. There was thus some similarity in the spectrum of streptomycetes, abundant in such communities and those found earlier to be able to interact in a distinctive way with Chlorella under experimental conditions. The majority of streptomycete strains (25 out of 30 checked) isolated from algo-bacterial communities were shown in the laboratory to be able to form aggregates on mixing with at least one strain of green algae, isolated form the same community. Some strains (11 out of 30) were able to interact in this way with either of the 3 strains of algae (Pleurococcus sp., Chlorella sp. 1, Chlorella sp. 2) tested. Given the experimental evidence available, it would seem premature to speculate on the possible nature, origin and significance of algo-actinomycete associations in general. In looking for an appropriate term for depicting even the most advanced cases of interactions supposed to be possible, at this time one should probably be content with the term "protocooperation" (McInerney, 1986), as used in the least restrictive way. Metabolic interactions between actinomycetes and algae emerge as an area worthy of future studies. It is hoped, however, that at least one simple point has been made clear in the foregoing, and that is that success of such studies would in a significant measure depend on the appropriate selection of partners. References Ahmadjan, V. (1980). Separation and artificial synthesis of lichens. In: Cook, C.B. et al. (eds.). Cellular Interactions in Symbiosis and Parasitism. Ohio Univ. Press, Columbus, pp. 3-29 Aleshina, E.N. & L.N.Makarovskaya (1956). Actinomycetes as antagonists. Mikrobiologia, 24:309-314 (in Russian) Beaman, B.L. (1981). Mechanisms of pathogenesis and host resistance to the actinomycetes. Zbl. Bakt. Suppl., 11:209-220 Beeking, J.H. (1981). The genus Frankia. In: Starr, M.P. et al. (eds.) The Prokaryotes. Handbook on Habitats, Isolation and Identification of Bacteria. Springer Verlag, Berlin, vol. 2, pp. 1191-2003 Belik, K., M.Herold & J.Doskocil (1957). New methods of biosynthetic production of antibiotics. 1. Biosynthesis of chlortetracycline without the maintenance of aseptic conditions. Folia Biol., 5:229-332 Benson, D.R. (1988). The genus Frankia: actinomycete- symbionts of plants. Microbiol. Sci., 5:9-12 Berry, A.M. (1986). Cellular aspects of root nodule establishment in Frankia symbiosis. In: Kosuge, T. & E.W.Nester (eds.). Plant-Microbe Interactions. Molecular and Genetic Perspectives. McMillan Publ. Co., New York, vol. 2, pp. 194-214 Buch, A. (1956). Isolation of microbial product affecting the synthesis of tetraene antibiotics by a strain of Streptomyces aureus. M.S. Thesis, Rutgers Univ., 1:92 Chaboud, A. & M. Lalonde (1983). Lectin binding on surfaces of Frankia strains. Can. J. Bot., 61:2889-2897 Cisar, J.O. (1986). Fimbrial lectins of the oral Actinomyces. In: Mirelman, D. (ed.) Microbial Lectins and Agglutinins: Properties and Biological Activity. Wiley Interscience, New York, pp. 183-195 Cook, R. (1977). The Biology of Symbiotic Fungi. Wiley & Sons, London, 389 pp. Craveri, R. (1956). Isolation and biological study of actinomycin. Rev. Biol., 43:239-244 Culberson, C.F., W.L.Culberson & A,Johnson (1984). Secondary compounds produced exclusively by lichens. In: Laskin, A.I. & H.A.Lechevalier (eds.) Handbook of Microbiology. CRC Press, Boca Raton, Fla., 2nd ed., vol. 5, pp. 793- 833 Dondero, N.C. & T. Scotti (1957). Excretion by streptomycetes of factors causing formation of aerial hyphae by old cultures. J. Bacteriol., 73: 584-585 Egorov, N.S., O.Z.Popova, M.V. Bitteeva, V.G. Bulgakova & K.Gofman (1960). Influence of bacterial metabolites on growth and antibiotic properities of some actinomycetes. Mikrobiologia, 29:269-275 (in Russian) Egorov, N.S. & N.S.Landau (1982). Biosynthesis of biologically active compounds by mixed cultures of microorganisms. Prikl. Mikrobiol. Biokhim., 18:835-849 (in Russian) Egorov, N.S. & N.S.Landau (1986). Influence of metabolic products on the character of populational interrelations in artificial ecosystems. In: Ecological Role of Microbial Metabolites. Moscow State University, Moscow, pp. 178-200 (in Russian) Fujita, Y., K.Oishi & K.Aida (1972). Hemagglutination by culture broth of actinomycctes and Asperigillus. J. Gen.Appl. Microbiol., 18: 73-75 Gause, G.F.T.P.Preobrazhenskaya, M.A. Sveshnikova, L.P.Terekhova, & T.S.Maksimova (1983). Determinative Mannual of Actinomycetes. Nauka, Moskow, 246 pp. (in Russian) Gitelson, I.I., N.S.Manukovskii & N.M.Pankova (1981). Microbiological Problems of Closed Ecological Systems. Nauka, Novosibirsk, 197 pp. (in Russian) Goodfellow, M. & S.T.Williams (1983). Ecology of actinomycetes. Ann. Rev. Microbiol., 57: 189-216 Gromov, B.V. The collection of algae cultures. Trud. Petergofsk. Biol. Inst. LGU, 19:149-154 (in Russian) Grossbard, E. (1955). Stimulation of pigmentation and sporulation in fungi by metabolites of actinomycetes. J. gen.Microbiol., 12: vi Kameyama, T.F.Ishikama, K.Oishi & K. Aida (1982). Comparison of l-fucose-specific lectins produced by six strains of Streptomyces. Agric. Biol. Chem., 46:523-528 Kozlova, A.N., G.V.Komarova, G.I.El-Registan & N.A.Krasilnikov (1971). Studies of metabolites of actinomycetes influencing the sexual function of Phycomyces blakesleeanus. Doklady AN SSSR, 200:230-232 (in Russian) Krasilnikov, N.A.A.I.Korenyako, N.I.Nikitina & G.K.Skryabin (1951). Intra- and interspecific interrelations and principles of species discrimination in microbes-antagonists. Izv.AN SSSR, Ser. Biolog., n. 4:66-80 (in Russian) Kuznetsova, N.A. (1974). Mixed cultivation of the mycoheptin producer with various microorganisms. Antibiotiki, 19:122-123 (in Russian) Kuznetsova, O.S., E.P.Yakovleva, T.P.Efimova & V.A.Tsyganov (1984). Effect of yeast rnetabolites on biosynthesis of levorin, levoristatin and fatty acids by the culture of Streptornyces levoris. Antibiotiki, 7:483-487 (in Russian) Lazo, W.K. (1964). An experimental association between Chlorella xanthella and Streptomyces. Amer. J. Bot., 51:678-679 Lazo, W.K. (1966). An experimcntal association between Chlorella xanthella and Streptomyces. Amer. J. Bot., 53:105-107 Likhacheva, A.A., G.M.Zenova & L.V.Kalakoutskii (1987). Interaction of streptomycetes and algae in mixed cultures. Mikrobiologia, 56: 309-313 (in Russian) Locci, R. & J.R.Locci (1975). On the alleged sexual cycle of a Streptomyces. Ann.Microbiol., 25:47-51 Maki, J.S. & R.Mitchell (1986). The function of lectins in interactions among marine bacteria, invertebrates, and algae. In: Mirelman, D. (ed.) Microbial Lectins and Agglutinins: Properties and Biological Activity. Wiley Interscience, New York, pp. 409425 McInerney, M.J. (1986). Transient and persistent associations among prokaryotes. In: Poindexter, J.S. & E.R.Leadbetter (eds.). Bacteria in Nature. Vol. 2. Methods and Special Applications in Bacterial Ecology. Plenum Press, New York, pp. 293-338 Millbank, I.W. (1976). Aspects of nitrogen metabolism in lichens. In: Brown, D.H., D.L.Hawksworth & R.H.Raily (cds.) Lichenology: Progress and Problems. Academic Press, London, pp. 441456 Mirelman, D. (ed.). (1986). Microbial Lectins and Agglulinins: Properties and Biological Activity. Wiley Interscience, New York Nakamura, H. (1963). Studies on the eco-system of Chlorella. Jap. Soc. Plant Physiol., p. 197 (Cited in: Vela, G.R., & C.N.Guerra, 1966) Ohshima, M., N.Ishizaki, A.Handa & Y.Tonooka (1983). Cultural conditions for production of neopurpuratin, a purplish-red pigment, by mixed cultures of Streptomyces purpuratus with Bacillus sp. J. Ferment. Technol., 61:31-36 Parker, B.C. & N.C.Bold (1961). Biotic relationships of soil algae and other microorganisms. Amer. J. Bot., 143:185- 197 Peveling, E. (1987). Lichens. In: Henis, Y. (ed.) Survival and Dormancy in Microorganisms. Wiley Interscience, New York, pp. 229-266 Polsinelli, M. & P.G.Mazza (1984). Use of membrane filters for selective isolation of actinomycetes from soil. FEMS Microbiol. Lett., 22: 79-83 Preobrazhenskaya, T.P., L.P.Terekhova, A. N. Kozlova & S.A.Egorova (1975). Production by Actinomyces malachitospinus of a physiologically active substance stimulating zygote formation in Mucorales. Mikrobiologia, 44: 641-644 (in Russian) Rehm, H.J. (1953). Untersuchungen uber das Verhalten von Pilzen und Streptomyceten in Mischkultur. Zbl. Bakt. Parasitenk. Inf. Hyg., Abt. 2, 107:418431 Rehm, H.J. (1959). Untersuchungen uber das Verhalten von Aspergillus niger und einem Streptomyces albus-Stamm in Mischkultur. Zbl. Bakt. Parasitenk. Inf. Hyg., Abt. 2, 112: 384402 Robinson, K. (1968). The use of cell wall analysis and gel electrophoresis for the identification of coryneform bacteria. In: Identification Methods for Microbiologists. Soc. Appl.Bacter. Techn.Ser., N. 2, part B, pp. 85-93 Smith, D.C. & A.E.Douglas (1987). The Biology of Symbiosis.Academic Press, London Stegeman, W.J. & J.K.Horber (1975). Induction of synthesis of bacterial protein by excretory product of the alga Chlamydomonas reinhardtii y-l. Nature, 257:244- 246 Tamija, H. & K.Shibata (1953). Effects of diurnally intermittent illumination on the growth and some cellular characteristics of Chlorella. In:Algal Culture from Laboratory to Pilot Plant. Carnegie Inst. Publ. 600, Washington, pp. 76-79 Tresner, H.D., J.A.Hayes & D.B.Borders (1971). Production of a naphthoquinone pigment by a species of Streptoverticillium and its accumulation by a streptomycete. Appl.Microbiol., 21:562-536 Vadjunina, A.F. & Z.A.Korchagina (1986). Methods of Study of Soil Physical Properties. Agropromizdat, Moscow, 415 pp. (in Russian) Vela, G.R. & C.N.Guerra (1966). On the nature of mixed cultures of Chlorella pyrenoidosa TX 71105 and various bacteria. J. gen. Microbiol., 42:123-131 Waksman, S.A. (1959). The Actinomycetes. Nature, Occurence and Activities. The Williams & Wilkins Co., Baltimore Williams, S.T., M.Goodfellow, G.Alderson, E.M.H.Wellington, P.H.A.Sneath & M.J. Sackin (1983). Numerical classification of Streptomyces and related genera. J.gen. Microbiol., 129:1743-1814 Yakovleva, E.P. (1983). Mixed cultivation of producers of biologically active substances with other microorganisms. Prikl.Mikrobiol.Biokhim., 29:330-346 (in Russian) Yakovleva, E.P. & E.D.Kuzmina (1985). Biosynthesis of lytic enzymes during the mixed cultivation of actinomycetes and yeasts. Antibiotiki, 12:887-893 (in Russian) Yakovleva, E.P., O.S.Kuznetsova & V.A.Tsyganov (1981). Excretion and some properties of the biostimulant formed by yeast-like fungi and its influence on biosynthesis of levorin. Antibiotiki, 26:327-332 (in Russian) Yakovleva, E.P., E.N.Sololova & V.A.Tsyganov (1972). Influence of conditions of joint cultivation of Actinomyces levoris with yeastlike fungi on levorin formation. Antibiotiki, 11: 981-975 (in Russian) Zenova, G.M., A.A.Likhacheva, S.O.Suetin & L.V.Kalakoutskii (1980). Mixed culture of an actinomycete and an alga. Biol. Nauki, 5:73-78 (in Russian) Zenova, G.M., G.M.Nikolaev, I.G.Sumarukova & L.V.Kalakoutskii (1986). Survivability of components of actinomycete-alga associations under conditions of low humidity. Biol.Nauki, 6:79-83 (in Russian) Zenova, G.M., E.B.Rydkina & L.V.Kalakoutskii (1983). Growth and antimicrobial activity of associations of actinomycete and green algae. Biol. Nauki, 3:81-85 (in Russian) Zenova, G.M., A.N.Efron, A.A.Likhacheva & L.V.Kalakoutskii (1988). Specific features of bacterial components of the algae - bacterial coenoses on outcrops of calcareous rocks. Vest. Moscow Univ. Ser.17, 1:44-48 (in Russian). Copyright 1990 C.E.T.A., The International Centre for Theoretical and Applied Ecology, Gorizia
The following images related to this document are available:Photo images[ac90006a.jpg] |
| |||||||||
{kind=link}