 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Donor-dependent Stimulation of Superoxide Output by Muramyl Dipeptide (MDP) in Human Granulocytes and Synergism of MDP with Lipopolysaccharide I. TARNOK and Zs. TARNOK Research Institute Borstel, Division of Bacteriology, D-2061 Borstel, FRG
Code Number: AC90007
Size of files
Text: 10.7K
Graphics: Line drawings (gif) - 23.8K Abstract. In the course of an investigation we observed: (1) a donor- dependent stimulation of the superoxide output by MDP and thus, this compound must be characterized as active in this respect. MDP activated mainly the second phase of the superoxide output; (2) no beneficial effect of a preincubation ("priming") with MDP prior to the addition of a second challenge (e.g., LPS). That means, preincubation with MDP did not lead to a higher superoxide production with endotoxin. Controls, preincubated with buffer instead of MDP showed the same superoxide output; thus, chemiluminescence counts might increase without a "priming" compound as well; (3) a synergism between MDP and LPS. The superoxide output was significantly higher when both substances were added simultaneously than when they were applied alone. The synthetic immunoadjuvant N-acetylmuramyl-L-alanyl-D isoglutamine (muramyl dipeptide, MDP) (Adam et al., 1974, 1975), related to the mycobacterial cell wall peptidoglycan, represents "the minimal structure responsible for many activities of bacteria, including immunoadjuvant effect" (Kaku et al., 1983). The peptidoglycan structure is present in the cell wall of the members of Actinomycetaceae as well and might act as immunoadjuvant. It is believed that MDP is unable to stimulate superoxide output in neutrophils (PMN) (Kaku et al, 1983, Guthrie et al., 1984, Wright and Mandell, 1986) and interacts with PMN only by means of preincubation ("priming"); that is "primed" PMN produce considerably higher amounts of superoxide in response to a subsequent second challenge than "nonprimed" cells. Similar results have been found in macrophages (Pabst and Johnston, 1980). Our own observations were in contrast to those of Kaku et al. (1983) and Wright and Mandell (1986). Firstly we obtained a considerable superoxide output in human PMN stimulated solely by MDP and secondly we were unable to obtain a beneficial effect from "priming" with MDP. Considering the fact that after a "priming" with MDP, PMN are exposed to two different substances simultaneously (MDP and e.g., LPS), it was justified to question whether MDP might act synergistically or antagonistically with other compounds. So far as we know, this possibility was not investigated up to now. In particular this paper deals with the following items: (a) superoxide output in human PMN stimulated directly by MDP;(b) superoxide output measured after preincubation ("priming") with MDP and subsequent challenge with lipopolysaccharide (endotoxin, LPS); (c) superoxide output following the simultaneous addition of MDP and LPS (antagonism or synergism) and (d) whether the superoxide output stimulation by MDP is donor-dependent. Materials and Methods Reagents. The following reagents were used: PBS (phosphate balanced salt solution according to Dulbecco), PBSAG (PBS supplemented with 0.1% human albumin and 1% glucose), lucigenin (1 mg lucigenin per ml in PBSAG), formylmethionyl-leucyl-phenylalanine (fMLP) (10 umol/l in PBSAG) and LPS (1 mg/ml in distilled water). PMN was separated according to West et al. (1981) from blood of healthy volunteers.
The suspension was adjusted to contain 20,000-40,000 cells in 20 ul PBSAG. Keeping it at room temperature, the suspension remained usable for 3-4 hours. An aliquot was kept at 0 C for a possible repetition of the measurements. The number of living cells was determined according to West et al. (1981). Staining for other blood cell types (eosinophils etc.) was carried out as well. Measurement of the direct superoxide output in the presence of MDP. 20 ul lucigenin solution, 40 ul PBSAG, 5 ug MDP (in 5 ul water) and 20 ul PMN suspension were pipetted into a polyethylene tube 0.8 cm in diameter, mixed well and chemoluminescence was measured for about 18 min in a Turner luminometer. Preincubation ('priming') with MDP. 20 ul lucigenin solution, 40 ul PBSAG, 5 ug MDP (in 5 ul water) and 20 ul PMN suspensions were incubated for different times in the luminometer; then, 1 ul LPS was added and chemoluminescence measured immediately. About 3 seconds were necessary to add the LPS and start the luminometer again. Synergism between MDP and LPS. The reaction mixture contained 20 ul lucigenin solution, 5 ug MDP (in 5 ul water), 40 ul PBSAG, 1 ul LPS and 20 ul PMN suspension. Chemoluminescence was measured immediately after the addition of PMN.
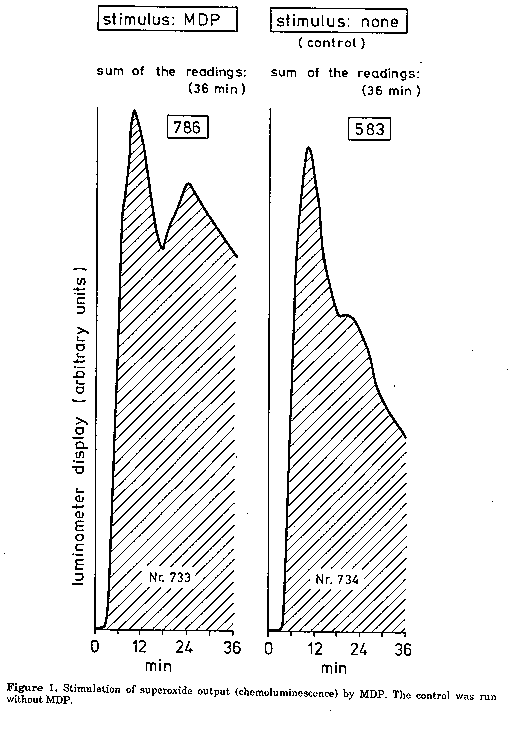
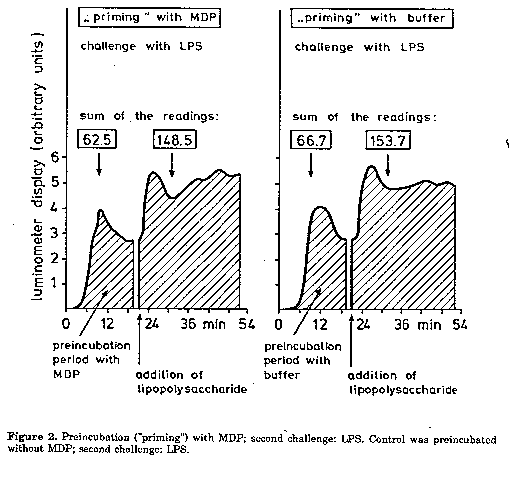
Effect of superoxide dismutase (SOD). In proof of superoxide formation, 15-25 units of SOD in 5 ul PBS were added to 60 ul reaction mixture as mentioned above; PMN were added last. SOD was purchased from Sigma (1 mg protein per ml; 3000-5000 units per mg protein). Catalase (CAT). The formation of hydrogen peroxide was controlled by using 10-30 units of CAT in 1 ul PBS (Sigma; 10,000-30,000 units per mg protein). For details see under SOD. Results and Discussion Stimulation of superoxide output by MDP. Our results are in disagreement with those published by Pabst and Johnston (1980), who reported negative results with MDP. MDP is unquestionably able to activate superoxide output in PMN (Fig. 1). However, the output depended on the donor in question: in some cell populations, we obtained an activation by MDP, in others it was not influenced. Preincubation ('priming') with MDP. This procedure did not show any beneficial effect (Fig. 2). Though a higher chemoluminescence could be observed with MDP after a priming period of 9 minutes, the control, "primed" just with buffer instead of MDP, showed nearly the same enhancement. This observation is in contradiction to those of Guthrie et al. (1984) and Wright and Mandell (1986) who observed a beneficial effect of the priming procedure. Table 1. Synergism between muramyl dipeptide (MDP) and lipopolysaccharide (LPS) in PMN; mean luminometer readings in the reaction mixtures (absolute values and percent of the control). Measurements without preincubation. Reaction mixture: 20 ul lucigenin solution, 38 ul Dulbecco's PBS supplemented with albumin and glucose, 20 ul PMN suspension. 5 ul MDP or LPS solution were used as indicated. Sum of luminometer readings for MDP and LPS added separately: 5086. Luminometer reading for MDP plus LPS added simultaneously: 5580; increase 9.7%.
Number PMN PMN PMN plus MDP PMN
and plus plus plus LPS plus
date MDP LPS simultaneously buffer
(5ug) (5ug) (5ug each) (control)
-------------------------------------------------------
733/26.4.89 796 ... ... ...
(136%)
736 " ... 4290 ... ...
(736%)
735 " ... ... 5580 ...
(957%)
734 " ... ... ... 583
(100%)
Synergism between MDP and LPS. It seemed to depend on the donor in question whether MDP could act synergistically or not. In PMN populations reacting well in this test, a considerably higher superoxide output was obtained when LPS and MDP were added simultaneously (Table 1). As for the kinetics of the enhanced superoxide formation, mainly the second phase of the usually biphasic output was activated; this corresponds to the presence of extracellular superoxide. The first phase (intracellular superoxide formation) remained unchanged or showed a certain degree of inhibition in comparison to the control. Tentatively we attribute this phenomenon to synergism; additional investigations will be carried out. Superoxide dismutase in the reaction mixture caused a strong decrease in the chemoluminescence providing evidence of superoxide formation; catalase was ineffective indicating that hydrogen peroxide was not involved. Acknowledgments. The authors are very much indebted to Dr.H.Brade for providing the LPS preparations and to Mrs.I.Hensel for isolating the granulocytes. References Adam, A., R.Ciorbaru, F.Ellouz, J.F.Petit & E. Lederer (1974). Adjuvant activity of monomeric bacterial cell wall peptidoglycans. Biochem. Biophys. Res. Comm., 56:561- 567 Adam, A., F-F. Ellouz, R.Ciorbaru, J.F. Petit & E. Lederer (1975). Peptidoglycan adjuvants: minimal structure required for activity. Z. Immunitatsforsch., 149:341-348 Guthrie, L.A., L.C.McPhail, P.M.Henson & R.B.Johnston (1984). Priming of neutrophils for enhanced release of oxygen metabolites by bacterial lipopolysaccharide. J.Exp. Med., 160: 1656-1671 Kaku, M., K.Yagawa, S.Nagao & A.Tanaka (1983). Enhanced superoxide artion release from phagocytes by muramy] dipeptide of lipopolysaccharide. Infect. Immun., 39:559-564 Pabst, M.J. & R.B Johnston (1980). Increased production of superoxide anion by macrophages exposed in vitro to muramyl dipeptide or lipopolysaccharide. J. Exp. Med., 151:101- 114 West, M.-Y., D.Sinclair & P.Southwell-Keely (1981). On the stoichiometry of oxygen metabolism in polymorphonuclear leukocytes. Biochem.Biophys. Res. Comm., 100:212-218 Wright, G.G. & G.L.Mandell (1986). Anthrax toxin blocks priming of neutrophils by lipopolysaccharide and by muramyl dipeptide. J. Exp. Med., 164: 1700-1709. Copyright 1990 C.E.T.A., The International Centre for Theoretical and Applied Ecology, Gorizia
The following images related to this document are available:Line drawing images[ac90007b.gif] [ac90007a.gif] |
| |||||||||
{kind=link}
{kind=link}