 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Use of the Skerman micromanipulator for isolating Actinomycetes in the wastewater field LINDA L. BLACKALL Animal Research Institute, Queensland, Australia.
Code Number: AC91003
Sizes of Files:
Text: 13.3K
Graphics: line drawings (gif) - 18.6K
Abstract. In the course of an investigation into foaming in activated sludge sewage treatment plants, a micromanipulation method for the isolation of the predominant foaming actinomycetes was employed. The Skerman Micromanipulator was the best method of confidently and quickly (pure cultures in 5-10 min) isolating the poorly competitive, slow growing, morphologically distinctive nocardioforms that were observed to comprise the bulk of the foam. One newly described foaming actinomycete (Nocardia pinensis), has never been isolated by any other method. Nocardioforms from many regions of the world have been isolated using this technique. The activated sludge sewage treatment process is commonly used for the treatment of domestic sewage. The sewage is first treated aerobically followed by sedimentation to separate the purified liquid from the biological sludge (Winkler and Thomas, 1984). As well as the desirable organisms being selected for in the activated sludge process, undesirable ones also compete well. One example of a biological problem in activated sludge plants is the formation of a large stable foam on the surface of the aeration tank section. When this foam is examined microscopically, it is often predominantly composed of Gram-positive branching filamentous bacteria (Blackall, 1990; Soddell and Seviour, 1990). The actino- mycetes associated with foaming in activated sludge plants are Nocardia amarae (Lechevalier and Lechevalier, 1974; Blackall, 1990), Nocardia pinensis (Blackall et al., 1989; Blackall, 1990), Rhodococcus rhodochrous (Lemmer and Kroppenstedt, 1984), and Tsukamurella paurometabolum (Lemmer and Kroppenstedt, 1984; Collins et al., 1988). The amount of foam biomass present on aeration tank surfaces emphasises the competitive ability of these nocardioform actinomycetes in their natural environment. Their numerical dominance is dramatically lowered in the laboratory when dilution and spread plating methods are used for their isolation. Indeed, if the predominant foaming actinomycete is N. pinensis, no colonies are recovered if this method of isolation is exploited. The foaming actinomycetes are morphologically distinctive, numerically dominant in the samples, poor competitors in batch conditions in the laboratory and as such are prime candidates for isolation by micromanipulation. Materials and methods Media.
Bacterial mycelium was micromanipulated to either yeast glucose agar (YG) containing (per litre), 10 g yeast extract, 10 g glucose and 15 g agar, to tryptone yeast extract agar (TYG) containing (per litre), 3 g tryptone, 5 g yeast extract, 5 g glucose and 15 g agar or to activated sludge agar (AS). The pH of all media was adjusted to 7.0 and YG and TYG were autoclaved at 121 C for 15 min. AS was prepared by crudely filtering mixed liquor from an activated sludge plant through absorbent cotton wool. Agar (1.5%) was added to the filtrate followed by autoclaving at 108 C for 30 min. Identification of isolates was done by previously reported methods (Blackall, 1987; Blackall et al., 1989). Samples. These were either foam or mixed liquor from activated sludge sewage treatment plants.
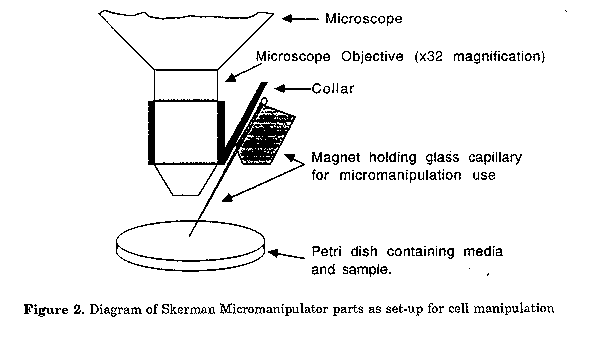
Figure 2. Diagram of Skerman Micromanipulator parts as set-up for cell manipulation
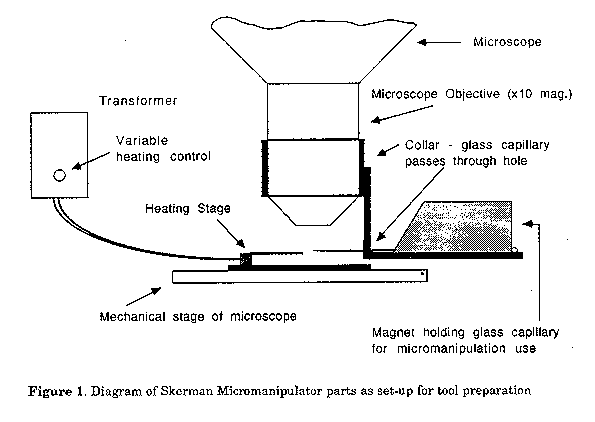
Activated sludge samples were serially diluted in distilled water and vortexed on high speed for 2 min. The liquid (0.1 ml) was drop inoculated onto the surface of the media listed above, spread with a spreader and incubated at 28 C. Presumptive nocardioform microcolonies were located with the assistance of phase contrast microscopy. Micromanipulation. The Skerman Micromanipulator (Skerman, 1968) was used. A Leitz Wetzlar, Phaco 1, 32x magnification (170/-, L 32/0.40) objective on an Olympus CHA microscope fitted with a phase contrast turret condenser was used for manipulation. The con- densor phase plate for the Olympus 20x magnification objective was used with the Leitz objective as the phase rings gave the closest fit. The operation of the micromanipulator will be briefly described. There are two parts to micromanipulation - tool preparation (Fig. 1) and use of the tools for micromanipulation of cells (Fig. 2).
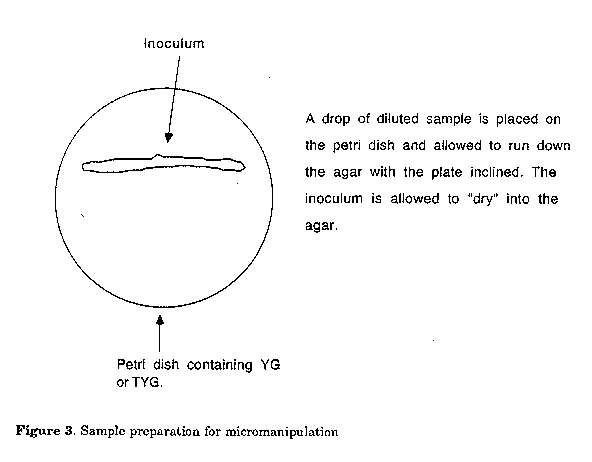
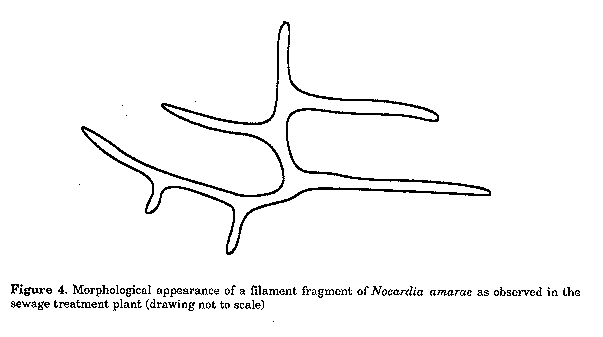
A drop of diluted sample is placed on the petri dish and allowed to run down the agar with the plate inclined. The inoculum is allowed to "dry" into the agar. Figure 4. Morphological appearance of a filament fragment of Nocardia amarae as observed in the sewage treatment plant (drawing not to scale)
Microtools (knobs, loops, hooks) are prepared from glass capillaries using a microforge comprised of a regulated power supply, a heating stage and a microforge toolcarrier. The glass capillary is fixed into a magnet that is located on a collar that fits neatly around the barrel of the objective. A xlO magnification objective is generally used for tool preparation. Under the microscope, the glass capillary is moulded to the desired shape, using the forge at the appropriate temperature. The microtool glass capillary is fixed into a magnet which facilitates movement from the preparation collar to the manipulation collar.
Tool Use. The microtool used for manipulation, is located on a collar that fits neatly around the barrel of the 32x objec- tive. The sample is appropriately diluted and inoculated onto a petri dish containing either YG or TYG (Fig. 3). This plate is placed on the mechanical stage of the microscope. During manipulation, the tool remains stationary, and the plate containing the sample from which cells are manipulated, is moved on the microscope stage. Once the target cells are microscopically located, they are moved away from the inoculum area with the microtool and this area is marked using the microtool. Using a plate microscope, the marked area can be excised with a sterile scalpel and inoculated to either solid or liquid media. To maintain "aseptic" conditions during manipulation, the microscope can be covered with a clear plastic bag containing holes for the protruding eyepieces.
Manipulated cells were inoculated to either AS, YG or TYG and incubated at 28 C. The plates were regularly examined by phase contrast microscopy employing the "sterile" plastic bag micro- scope cover to reduce possible contamination. Results Using conventional dilution and spread plating methods and YG media, a range of nocardioform isolates were acquired. A small percentage of them were identified as Nocardia amarae, however, the vast majority were unidentifiable. None were identified as Nocardia pinensis. After micromanipulation to YG was used to isolate the predominant nocardioforms from the samples, only N. amarae was isolated. In the sewage treatment plant, the filaments produced branches perpendicular to the main stem (Fig. 4). Cardinal resolving properties of N. amarae are: rifampicin sensitivity (20 ug ml^-1), isonicotinic acid hydrazine resistance (200 ug ml^-1), degradation of hypoxanthine, xanthine, tyrosine and casein, lysozyme sensitivity (50 ug ml^-1), lypolysis of Tween aesculin hydrolysis, presence of polyphosphate and sudanophilic inclusions and non-acid fastness.
It was observed that the morphology of the filaments in the activated sludge foams sometimes differed from the con- ventional perpendicular branching nocardioforms. They were like a pine tree in cellular morphology i.e. the branches were at a more acute angle to the primary mycelium (Fig. 5).
Using micromanipulation, nocardioform actinomycetes have been isolated from sewage treatment plants in Australia, the US, and Italy. Discussion The isolation of the prominent nocardioform actinomycetes from foams in activated sludge plants, has been dramatically improved by the introduction and use of the Skerman Micromanipulator. These organisms are prime candidates for this methodology. Micromanipulation is the only method where one can be quite confident that the organisms isolated are the ones that are wanted. The appearance of R. rhodochrous, T. paurometabolum, and N. amarae in activated sludge are all similar and nocardioform micro- colonies are all fairly similar in appearance. N. amarae is one of the more slow growing nocardias and nocardioform actinomycetes are common members of the activated sludge microflora. If other nocardioforms are present in activated sludge in lower numbers than N. amarae, the former could still be isolated because they could grow to larger colonies more quickly. The isolation of N.pinensis had eluded researchers until micromanipulation was used.
The physiological data (kinetic constants and substrate affinities) for N. amarae are being used to devise methods to control foaming in activated sludge. This can be achieved by altering the configuration of the plant and thus changing the pressures that select the flora. If the physiological data for the wrong organism is used to justify modification of the plant, then the control strategy may be unsuccessful. This underlines the importance of isolation of the correct foaming organism. The physiological data for N. pinensis is not yet available, however, the sewage treatment plant modifications that lead to the control of foaming caused by N. amarae may not control the growth and competitive advantages of N.pinensis The simplicity of the method, moderate cost of the equipment and ease with which operators can acquire the ability to use the micromanipulator, make this method of selective isolation an attractive one. References Blackall, L.L. (1987). Actinomycete scum problems in activated sludge plants. Ph.D. Thesis, University of Queensland, St. Lucia, Australia. Blackall, L.L. (1990). The microbiology of bulking and foaming in activated sludge. Water, 17: 4446 Blackall, L.L., J.H.Parlett, A.C.Hayward, D.E.Minnikin, P.F.Greenfield & A.E.Harbers (1989). Nocardia pinensis sp.nov., an actinomycete found in activated sludge foams in Australia. J. gen. Microbiol., 135:1547-1558 Collins, M.D., J.Smida, M.Dorsch & E.Stackebrandt (1988). Tsukamurella gen. nov. harboring Corynebacterium paurometabolum and Rhodococcus aurantiacus. Int. J. System. Bacteriol., 38:385-391 Lechevalier, M.P. & H.A. Lechevalier (1974). Nocardia amarae sp. nov., an actinomycete common in foaming activated sludge. Int. J. System. Bacteriol., 24:278-288 Lemmer, H. & R.M.Kroppenstedt (1984). Chemotaxonomy and physiology of some actinomycetes' isolated from scumming activated sludge. System.Appl. Microbiol., 5: 124-153 Skerman, V.B.D. (1968). A new type of micromanipulator and microforge. J.gen.Microbiol., 54:287-297 Soddell, J.A. & R.J.Seviour (1990). Microbiology of foaming in activated sludge plants. J.Appl. Bacteriol., 69: 145-176 Winkler, M.A. & A.Thomas (1984). Biological treatment of aqueous waste. In: Wiseman, A. (ed.). Topics in Enzyme and Fermentation Biotechnology. Ellis Horwood Ltd., Chichester, Vol 1, pp. 200-279. Copyright 1991 C.E.T.A., The International Centre for Theoretical and Applied Ecology, Gorizia The following images related to this document are available:Line drawing images[ac91003d.gif] [ac91003c.gif] [ac91003b.gif] [ac91003a.gif] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}