 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ultrastructural changes in the cells of the Streptomyces spectabilis 1011-10 strain during biosynthesis of streptovaricin
D.SPASSOVA, N.VESSELINOVA and R.GESHEVA Institute of Microbiology, Bulgarian Academy of Sciences, Sofia, Bulgaria
Code Number: AC91005
Sizes of Files:
Text: 17.5K
Graphics: line drawings (gif) - 3.5K
photographs (jpg) - 453.0K
Abstract. Ultrastructural changes in the cells of a highly active naturally selected strain of Streptomyces spectabilis, 1011-10, during biosynthesis of streptovaricin are reported. Ultrastructural peculiarities in the cells of the strain, such as large vacuoles, interruption of the cell wall in the vicinity of large vacuoles, release of globules of different size and ribosome aggregation were not observed in the parent strain of S.spectabilis with low antibiotic activity. Streptovaricins were among the first antibiotics discovered in the group of the ansamycins .(Rhuland et al., 1957; Siminoff et al., 1975). They inhibit the RNA synthesis of bacteria. It is known that Streptomyces spectabilis strains are producers of streptovaricin. In previous investigations we found that Streptomyces sp. 1000, isolated by us, belonged to this species (Vesselinova et al., 1990). As a result of natural selection, the highly active S.spectabilis strain 1011-10, a producer of streptovaricin, was obtained.
The present work is aimed at establishing the ultrastructural changes of the S.spectabilis 1011-10 strain during biosynthesis and streptovaricin production. Materials and methods Strains and conditions of cultivation. S.spectabilis, strain 1000, and its natu- rally selected strain 1011-10, were studied. Cultivation was carried out on a shaker, 220 rpm at 28 C, in flasks containing 50 ml of complex soya medium.
Antibiotic activity. This was determined by bioassay in a solid medium against a Bacillus subtilis test culture.
Electron microscopy. Electron microscopic observations of the actinomycete strains were carried out on cultures between 24 and 96 h of cultivation. Cells were fixed according to the method of Kellenberg et al.(1958), and embedded in Durcupan (Fluka). The ultrathin sections were obtained on a Reichert-Joung ultramicrotom and were stained with lead citrate (Reynolds, 1963). Observations were carried out on an Opton EM 10C electron microscope at accelerating voltage of 60-100kV.
Comparative electron-microscopic investigations of the highly active S. spectabilis 1011-10 strain and of the slightly active parent strain, S. spectabilis 1000, were carried out to study the ultrastructural changes taking place during the bioproduction of streptovaricin.
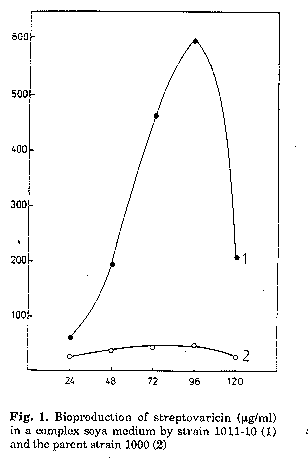
In parallel with ultrastructural investigations, the accumulation of streptovaricin in the culture medium was measured. As shown in Fig. 1, optimum antibiotic production by both strains occurred after 96h. The optimum streptovaricin production by the selected strain (595 ug/ml) was 14.5 times higher than that of the parent strain. The parent strain showed low activity during the entire course of its growth cycle.
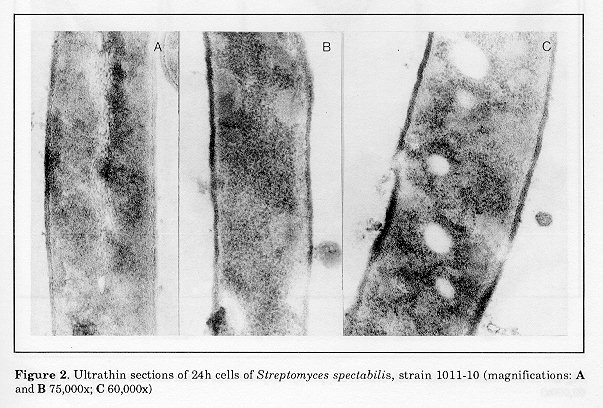
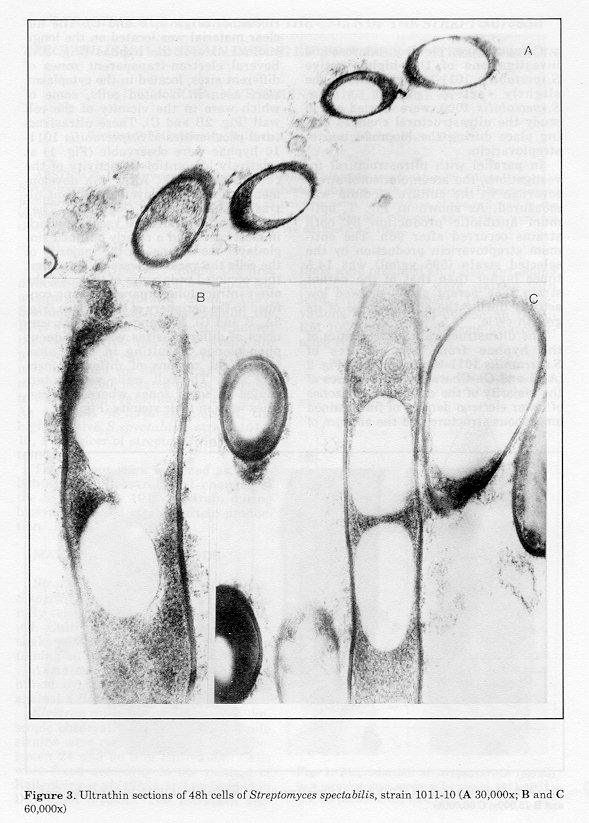
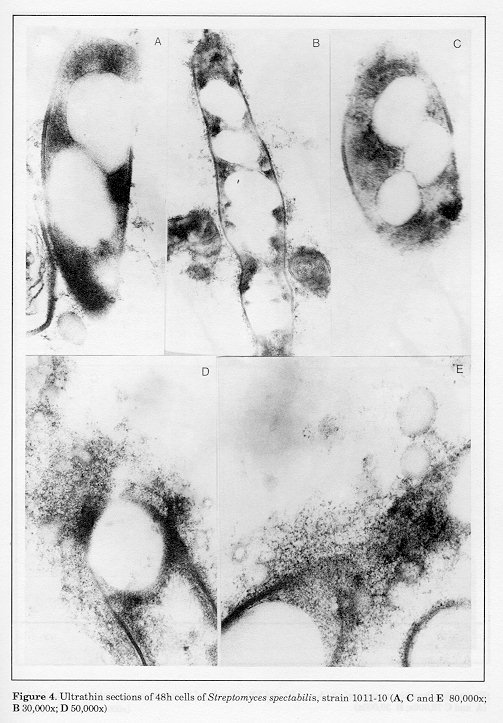
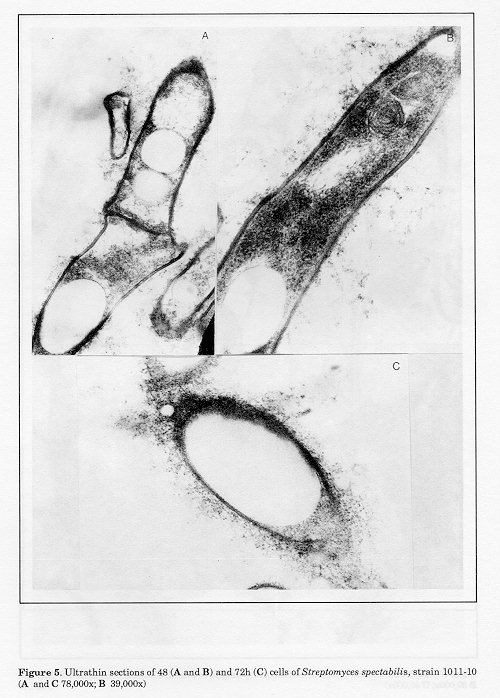
The ultrastructural characteristics of the hyphae from 24h cultures of S.spectabilis 1011-10 are shown in Fig. 2 (A, B and C). Characteristic features of the majority of the cells were the zones of lower electron density of fine-grained amorphous structure and the absence of ribosomes (Figs. 2, B and C). The nuclear material was located on the longitudinal axis of the hyphae (Fig. 2A). Several electron-transparent zones of different sizes, located in the cytoplasm, were seen in isolated cells, some of which were in the vicinity of the cell wall (Fig. 2B and C). These ultrastructural peculiarities of S. spectabilis 101110 hyphae were observable (Fig. 1) at relatively low antibiotic activity of the culture (72 ug/ml). After 48h development, all cells displayed significant ultrastructural changes (Figs. 3-5). They had one or several large vacuoles which in most cases had a clearly outlined sin- gle-layer membrane. In the majority of the cells the vacuoles were so large that they occupied wide zones of the cells, the other intracellular organelles being confined to the periphery of the hyphae (Fig. 3A, B and C). Merging of adjacent vacuoles of different sizes was a frequent phenomenon, resulting in wide electron-bright regions of different form (Fig. 4B). The cell walls were interrupted in some zones where the vacuoles were in their vicinity (Fig. 4A).
Figure 3. Ultrathin sections of 48h cells of Streptomyces spectabilis, strain 1011-10 (A 30,000x; B and C 60,000x) Figure 4. Ultrathin sections of 48h cells of Streptomyces spectabilis, strain 1011-10 (A, C and E 80,000x; B 30,000x; D
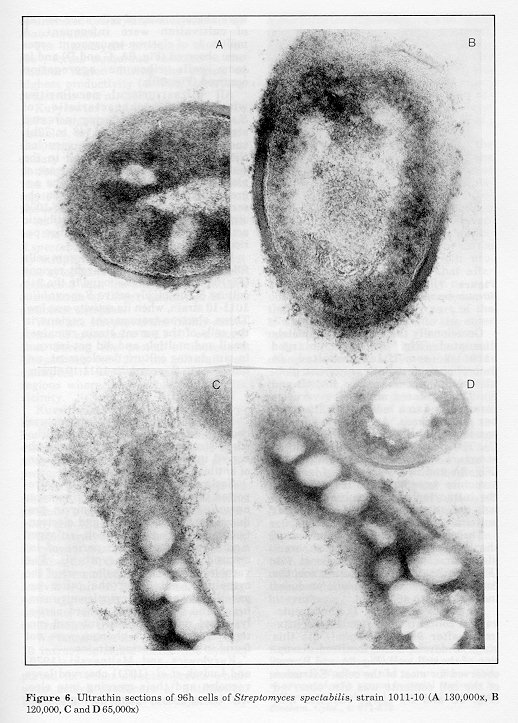
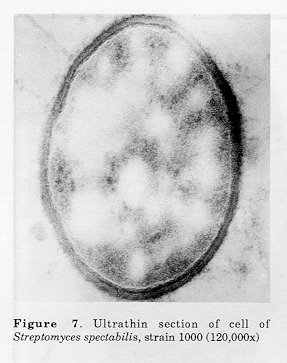
Figure 5. Ultrathin sections of 48 (A and B) and 72h (C) cells of Streptomyces spectabilis, strain 1011-10 (A and C 78,000x; B 39,000x) Figure 6. Ultrathin sections of 96h cells of Streptomyces spectabilis, strain 1011-10 (A 130,000x, B 120,000, C and D 65,000x) Figure 7. Ultrathin section of cell of Streptomyces spectabilis, strain 1000 (120,000x)
These ultrastructural peculiarities of the hyphae, observed after 48h, were also characteristic for the cells at 72h (Fig. 5C). At this stage of cultivation, the concentration of the antibiotic produced increased up to 463 ug/ml.
As can be seen (Fig. 1), the streptovaricin concentration reached its maximum after 96h (595 ug/ml). At this stage of development, localized disruption of the cell wall (Fig. 6A and B) was observed for most of the cells. Extrusion of the cell contents was also observed (Fig. 6C and D). Large vacuoles that were characteristic of cells after 48-72h of cultivation were infrequent. A multitude of electron-transparent zones were observed (Fig. 6A, C and D) and in some cells ribosome aggregation occurred (Fig. 6B). All ultrastructural peculiarities which were characteristic of S. spectabilis 1011-10 during increased streptovaricin biosynthesis (48 to 72h), such as the presence of large vacuoles, the disruption of the cell wall in the vicinity of large vacuoles, the release of globules of different sizes and the aggregation of ribosomes were not ob- served in the parent S. spectabilis 1000 strain. This strain showed low antibiotic activity (41 ug/ml) during the entire period of development (Fig. 1).
In the parent culture there were cells filled with electron-transparent regions (Fig. 6), similar to those found in the 24h culture of the highly-active S. spectabilis 1011-10 strain, when its activity was low. These electron-transparent regions in the cells of the parent strain remained small and multiple and did not increase in size during culture development, unlike those in S.spectabilis 1011-10 strain. Discussion Some of the observed ultrastructural changes during the intensive biosynthesis of streptovaricin have also been ob- served in other highly-active producers of antibiotics.
Saslavskaya and Shukov (1976) reported that the hyphae of "Actinomyces parvulus", an active actinomycin producer, contained vacuoles and electrontransparent regions which enlarged significantly during the period of increased antibiotic biosynthesis. The vacuoles occupied the major part of the cytoplasm, extruding the other organelles towards the cytoplasmic membrane and the cell wall, where partial lysis and release of their internal contents occurred. Such changes were not found in non producing strains.
Kurylowicz and Malinovski (1970) and Ludvik et al. (1971) observed large vacuoles and their merging into electron- transparent regions during periods of intensive tetracycline biosynthesis by S.aureofaciens.
The lincomycin producer, "A.roseolus", forms numerous vacuoles in the nuclear region during the period of highest productivity (48-72h), according to Listvinova et al. (1980).
Kurylowicz and Malinovski (1970, 1972) suggested a correlation between vacuoles, on one hand, and the production, accumulation and secretion of secondary metabolites, on the other. The antibiotic localization in these regions was considered as a means of separating it from potentially sensitive parts of the cells.
As we did not observe large vacuoles in the cells of the parent culture S. spectabilis 1000, which is a low-activity streptovaricin producer, we suggest that cytoplasmic vacualization is closely connected with the biosynthesis and accumulation of larger quantities of the antibiotic complex.
With the streptovaricin highly-active strain (S. spectabilis 1011-10) we found that during the period of increased biosynthesis (48-72h), the cell wall was disrupted in some zones, usually in the regions where the vacuoles were in its vicinity.
Kurylowicz and collaborators (Kurylowicz and Malinovski, 1970, 1972; Kurylowicz et al., 1975) observed cell wall disruption with highly-active actinomycete strains in the period of the production of given antibiotics.
Like us, some authors have observed ribosome aggregation in the antibiotic producer's cells: examples include actinomycin (Saslavskaya and Shukov, 1976), tetracycline (Ludvik et al., 1971; Kurylowicz et al., 1974; Saslavskaya and Shukov, 1976; Saslavskaya et al., 1977) and chrysomallin (Kuimova et al., 1978). Ludvik et al. (1971) assumed that ribosome aggregation was linked to the endogenous effect of high tetracycline concentration in the actinomycete cell.
We found that the S. spectabilis 101110 intracytoplasmic membrane system was not as well developed as that reported for other streptomycetes which were active producers of antibiotics. Examples include: tetracycline by S.aureofaciens (Kurylowicz and Malinovski, 1972; Kurylowicz et al., 1974), erythromyci.n by S.erythreus (Kurylowicz et al., 1974), candistin by S. noursei (Kurylowicz et al., 1974), viomycin by S. vinaceus (Kurylowicz et al., 1974), chrysomallin by "A.chrysomallus" (Kuimova et al., 1978) and flavomycin by "A.flavus" (Kulalaeva et al., 1976).
Saslavskaya et al. (1978) studied the special ultrastructural features of the active as well as of the inactive varieties of the producer of the ansamycin antibi- otic, rifamycin. Despite producers' belonging to another genus (Nocardia mediterranei), it was found that vacuoles decreased with increasing antibiotic activity, a fact established by us too during maximum streptovaricin pro- duction. It can be assumed that after 96h of cultivation, practically no vacuoles in the cells were observed due to the transport of the major part of the biosynthetized antibiotic into the outer medium.
In summary the ultrastructural changes in the cells of S.spectabilis 1011-10 strain during the optimum period of biosynthesis of streptovaricin (48-72h) were: vacuole enlargement, thinning out and disruption of parts of the cell wall in the vicinity of the vacuoles, release of globules of different size and aggregation of ribosomes. These were not observed in the hyphae of the parent strain which exhibited low antibiotic activity. These morphological changes correlate with the functional activity of S. spectabilis 1011-10 strain during the active biosynthesis phase. They were not typical for the 96h culture, when the antibiotic had been released into the culture medium. The changes described could be used as morphological parameters in the microbiological control of biosynthesis. References Kellenberg, E., A.Ryter & J.Sechard (1958). Electron microscope study of DNA containing plasms. II. Vegetative and mature phage DNA as compared with normal bacterial nucleoids in different physiological states. J. Biophys. Biochem. Cytol., 4:671-676 Kuimova, T.F., V.S.Soina, A.A. Sokolov & O.I. Artamonova (1978). Electron microscopic study of the structure of mycelium of Actinomyces chrysomallus producing the antibiotic chrysomallin in the process of submerged fermenta- tion. Mikrobiologiya, 47:745-749 Kulalaeva, Z.I., V.A.Poltorak, E.I.Zaichina, V.S.Tjurin & A.B.Silaev (1976). Vegetative mycelium of Actinomyces flavus by electron microscopy in the course of biosynthesis of flavofungin. Mikrobiologiya, 45:133-136 Kurylowicz, W. & K.Malinovski (1970). Electron microscopy of surface of two strains of Streptomyces aureofaciens during tetracycline biosynthesis. Acta Microbiol. Pol., Ser. B, 2: 223-228 Kurylowicz, W. & K.Malinovski (1972). Ultrastructure of the mycelium of Streptomyces aureofaciens in the course of biosynthesis of tetracycline. Postepy Hig. Med. Dosw., 26:563-569 Kurylowicz, W., W.Kurzatkowski, W.Woznicka, A.Paskiewicz & S.T.Williams (1974). Ultrastructure of selected strains of actinomycetes during biosynthesis of antibiotics. Post. Hig. Med. Dosw., 28:419-425 Kurylowicz, W., W.Kurzatkowski, S.T. Williams, W.Woznicka & A.Paskiewicz (1975). Atlas of Ultrastructure of Streptomyces in Course of Biosynthesis of Antibiotics. Polish Medical Publishers, Warsaw Listvinova, S.N., P.L.Saslavskaya, S.V.Dmitrieva & N.V.Prilova (1980). Effect of lincomycin on Actinomyces roseolus producing it during prolonged passage of liquid medium. Antibiotiki, 25:88-92 Ludvik, T, K.Mikulik & Z.Vanek (1971). Fine structure of Streptomyces aureofaciens producing tetracycline. Folia Microbiol. (Prague), 16: 479480 Reynolds, E.S. (1963). The use of lead citrate at high pH as an electron opaque stain in electron microscopy. J. Cell Bid., 17:208-212 Rubland, L.E., K.Stern and H.R.Reames (1957). Streptovaricin. III. In vivo studies in tuberculous mouse. Am. Rev. Tuberc. Pulm. Dis., 75:588-659 Saslavskaya, P.L. & V.T.Shukov (1976). Morphological changes in the mycelium of active and inactive variants of Actinomyces parvullus in the course of its growth and biosynthesis of actinomycin D. Mikrobiologiya, 45:1035-1039 Saslavskaya, P.L., V.G.Makarevich,& M.D. Slugina (1977). Morphological and functional investigations of growth of Actinomyces aureofaciens in conditions of regulated and non-regulated fermentation. Mikrobiologiya, 46: 283- 287 Saslavskaya, P.L., A.F.Teteriatnik, V.T. Shukov & N.D.Dzegilenko (1978). Electron microscopic investigation of active and inactive variants ofProactinomyces (Nocardia) mediterranei, an organism producing rifamycin B. Antibiotiki, 23:208-211 Siminoff, P., R.M.Smith, W.T.Sokolovski & G.M.Savage (1975). Streptovaricin. I. Discovery and biological activity. Am.Rev. Tuberc. Pulm. Dis., 75:576-583
Vesselinova, N., R.Gesheva & V.Ivanova (1990). Streptomyces sp. 1000 producing streptovaricin complex. Folia Microbiol. (Prague), (in press). Copyright 1991 C.E.T.A., The International Centre for Theoretical and Applied Ecology, Gorizia The following images related to this document are available:Photo images[ac91005d.jpg] [ac91005g.jpg] [ac91005b.jpg] [ac91005c.jpg] [ac91005f.jpg] [ac91005e.jpg]Line drawing images[ac91005a.gif] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}