 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Identification of some mycolic acid containing actinomycetes using fluorogenic probes based on 7-amino-4- methylcoumarin and 4-methyl-umbelliferone MICHAEL GOODFELLOW Department of Microbiology, The Medical School, Framlington Place, Newcastle upon Tyne, NE2 HH, U.K.
Code Number: AC91008
Size of Files:
Text: 25.2K
Graphics: line drawings (gif) - 11.1K
Identification is a branch of systematics. It is the act and
the result of determining whether a new isolate belongs to a
group derived from classification. The organism is given a
name by recognising that it is a member of a previously
defined taxon. In making an identification the unknown
organism is examined and its characters are compared with
those of characteristic organisms or descriptions of
characteristic organisms. Identification makes use of
weighted characters, that is, those which have been shown to
be significant in distinguishing one organism from another. In
this it differs markedly from classification, in which
unweighted characters are preferable.
There is no choice but to classify, name and identify. At a purely practical level, microbiologists need to know what organisms they are working with before they can pass on information about them. Sound classification is a prerequisite for stable nomenclature and accurate identification. Extensive taxonomic databases are invaluable for generating reliable identification schemes for separating organisms below the genus level. Numerical taxonomy provides such data on strains assigned to numerically defined clusters. The results of such studies are usually expressed as the number or percentage of strains within each duster which show a positive state for each of the features used to construct the classification. When classification is complete, databases can be searched for presumptive diagnostic characters. Those found to be reproducible can then be weighted and used in dichotomous keys, diagnostic tables and computer probability matrices. The application of this procedure to the identification of streptomycetes has been considered in detail by Williams et al.(l).
Large areas of microbiology are highly dependant on good identification. Some areas, such as hospital laboratories, are almost entirely concerned with identification and are collectively referred to as diagnostic microbiology. It is, however, not always appreciated that numerous other kinds of microbiological work, such as ecology and biotechnology studies, are severely hampered by the lack of good identification schemes. The choice of actinomycetes for industrial screening programmes, especially those with a low throughput, is essentially a problem of distinguishing between known actinomycetes and recognising new ones(2). This problem is partly historical the need to identify unknown actinomycetes having rarely been recognised as a relevant task in the industrial domain. Consequently, current protocols for the selective isolation of actinomycetes for screening pro- grammes owe as much to "custom and practice" as to any underlying scientific rationale. Identification of actinomycetes The identification of actinomycetes is essentially a two-fold process. Reliable criteria are needed to assign organisms to families and genera prior to the selection of diagnostic tests for identification to species and subspecific levels. Identification at generic and suprageneric ranks can usually be achieved using a combination of morphological and chemical criteria(3,4). In contrast, few reliable and well tested schemes are available for the differentiation of species. Those which are available involve considerably more effort than can easily be handled in most diagnostic and industrial screening laboratories.
Many methods have been recommended for the extraction and analysis of chemical markers(5). All these methods require little analytical chemical experience and can be performed relatively quickly with a minimum of equipment. The type of analyses needed to separate the major suprageneric groups of actinomycetes are outlined in Figure 1.
Enzymes as taxonomic markers One of the most outstanding properties of enzymes is their specificity. Some enzymes have almost absolute specificity for a given substrate and do not attack even closely related molecules; others are less so and act on a. class of compounds. Aspartase is an enzyme showing high specificity. It catalyses the reversible addition of ammonia to the double bond of fumarate to form L-aspartate. Glycosidases, peptidases and phosphatases show more general activity. This specificity of enzymes combined with their ability to catalyse reactions of substrates at extremely low concentrations has been used for taxonomic purposes ever since it became apparent that enzymes were discontinuously distributed amongst bacteria.
Most of the early enzyme tests used for bacterial identification were developed and applied empirically; their underlying biochemical basis became apparent only at a later date. Many of the chemical methods used to derive biochemical fingerprints of bacteria were lengthy, laborious and insensitive. The need for sensitivity and shorter incubation times led to the development of improved procedures for biochemical profiling. Chromogenic substrates were found to be highly reactive and sufficiently sensitive to demonstrate a range of enzyme activities in suspensions of non-growing bacterial cultures(10-12). The interpretation of results obtained with such tests is not complicated by the effects .of the multiple reactions that can occur in complex nutrient media containing the substrate(11).
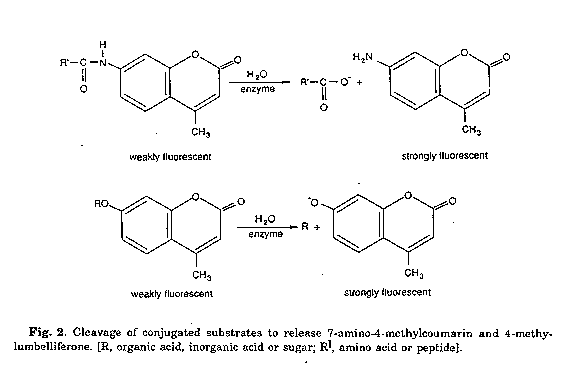
Fluorescence is a much more sensitive method for detecting enzyme activity. Colorimetric tests may be adapted for fluorometry by adding a fluorophore with a spectrum that is quenched by a product of the enzymic reaction or by using fluorescent indicators, notably 7-amino-4-methylcoumarin (7AMC) and 4-methylumbelliferone (4MU). Conjugated derivatives of these molecules can be purchased from Bachem Feinchemi- kalien A.G. (Bubendorf, Switzerland), Koch-Light Ltd. (Haverhill, Suffolk, U.K.), Novabiochem (U.K.) Ltd. (Uni- versity Boulevard, Nottingham, U.K.) and Sigma Chemical Co., Ltd. (Poole, Dorset, U.K.).
In all cases the conjugated substrates are cleaved by the relevant hydrolytic enzyme and the parent molecule released (Fig. 2). The parent molecules are intensely fluorescent in the visible region of the electromagnetic spectrum whereas the corresponding derivatives are only weakly fluorescent in this region. A restricted range of 4MU-linked substrates have been used in the classification and identification of diverse bac- teria, including mycobacteria(17,18), renibacteria(19) and streptomycetes(20). Similarly, representative rhodococci were characterised using peptide hydrolase substrates based on 7-amino-4-methylcoumarin(21). The analysis procedure(22) is very simple and suitable for the examination of large numbers of samples taken directly from selective isolation plates. Actinomycete inocula are applied, together with a drop of the conjugated (non-fluorescing) substrate, to the surface of Whatman No. 3 filter paper. Incubation is for 20 minutes at 37 C; a drop of saturated sodium bicarbonate is added to incubated 4MU preparations to enhance fluorescence.
--------------------------------------------------------------
Table 1. Enzyme tests differentiating the species of the
genus Rhodococcus
--------------------------------------------------------------
Species* a b c d e f g h i j k l m
No. of Strains 2 1 4 10 5 7 4 8 1 6 4 5 5
--------------------------------------------------------------
Number of strains giving positive result
A.Cleavage of 7-amino-
4-methylcoumarin (-7 AMC) substrates
Endopeptidases:
Z-Phenylalamine-
arginine-7AMC 2 1 4 0 0 7 4 7 1 6 4 0 0
Exopeptidases:
beta-Alanine-7AMC 0 0 4 10 5 0 2 8 1 6 4 5 5
iso-Leucine-7AMC 2 1 0 5 5 7 4 8 1 5 4 5 0
Pyroglutamate-7AMC 0 0 0 0 0 7 0 8 0 0 0 0 0
Threonine-7AMC 2 1 4 1 5 7 0 8 1 0 4 5 4
Valine-7AMC 2 0 0 3 5 7 4 8 0 5 4 5 5
B. Cleavage of 4-methylumbelliferone (4MU-) substrates
Glycosides:
4MU-beta-D-
Cellobiopyranoside 1 0 0 0 5 0 4 0 0 0 0 0 5
4MU-beta-D-
Galactopyranoside 2 0 0 10 0 0 4 0 1 0 0 0 0
4MU-beta-D-
Glucopyranoside 2 1 4 10 5 7 4 0 0 0 0 5 5
4MU-alpha-D-
Mannopyranoside 0 1 0 0 0 0 0 0 0 0 0 0 5
4MU-beta-D-
Mannopyranoside 0 0 0 0 5 0 4 0 0 0 0 0 0
4MU-beta-D-
Xylopyranoside 2 1 0 10 5 0 4 0 0 0 0 5 5
Inorganic esters:
4MU-Bis-phosphate 2 1 1 10 5 0 4 7 0 0 0 5 5
Organic ester:
4MU-Lignocerate 0 0 1 4 5 7 0 8 0 0 4 5 1
Abbreviation: Z, the N-terminal carbobenzoxy-group of
endopeptidase substrates.
*a= R. aichiensis h= R. luteus
b= R. chlorophenolicus i= R. marinonascens
c= R. coprophilus j= R. maris
d= R. equi k= R. rhodnii
e= R. erythropolis l= R. rhodochrous
f= R. fasciens m= R. ruber
g= R. globerulus
--------------------------------------------------------------
Positive reactions are recorded when a light-blue fluorescence is observed under a strong UV (366nm) light.
It can be concluded from the preliminary studies outlined above that fluorogenic probes prepared from 7-amino-4- methylcoumarin and 4-methylumbelliferone provide a simple, rapid and inexpensive way of detecting specific enzymes in small amounts of whole actinomycetes. The pilot study on the rhodococci (21) was extended by examining many strains for a diverse range of 7AMC- and 4MU-conjugated substrates in order to determine the full potential of such rapid tests in the characterisation and identification of Gordona, Rhodococcus and Tsukamurella species(I3,14). Identification of Gordona, Rhodococcus and Tsukamurella species The taxonomy of actinomycetes containing mycolic acids, high molecular weight 3-hydroxy fatty acids with a long alkyl branch in the 2 position, has undergone marked revision given the development and application of chemical, numerical and molecular systematic methods(23,24). Mycolic acid containing actinomycetes have many properties in common, form a distinct suprageneric group, and can be readily assigned to the genera Corynebacterium, Gordona, Mycobacterium, Nocardia, Rhodococcus and Tsukamurella using a combination of chemical and morphological properties(24).
--------------------------------------------------------------
Table 2. Enzyme tests differentiating the species of the
genera Gordona and Tsukamurella
--------------------------------------------------------------
Species* a b c d e f g
No. of Strains 4 6 2 8 4 14 9
--------------------------------------------------------------
Number of strains giving positive results
A.Cleavage of 7-amino-4-methylcoumarin (-7 AMC) substrates
Endopeptidases:
Z-Glycine-glycine-proline-7AMC 0 0 0 8 4 13 0
Z-Phenylalanine-arginine-7AMC 0 6 2 8 4 13 9
Exopeptidases:
D-Alanine-7AMC 0 6 0 8 4 0 9
iso-Leucine-7AMC 4 0 2 0 0 9 9
Pyroglutamate-7AMC 0 0 0 0 4 9 0
B.Cleavage of 4-methylumbelliferone (4MU-) substrates
Glycosides:
4MU-alpha-L-Arabinofuranoside 0 3 2 0 0 13 0
4MU-beta-L-Arabinofuranoside 0 1 0 0 0 1 9
4MU-alpha-D-Mannopyranoside 0 0 0 0 0 14 0
4MU-beta-D-Mannopyranoside 0 6 2 0 0 13 0
Inorganic esters:
4MU-Bis-phosphate. pH 5.2 0 3 2 8 4 14 9
4MU-Pyrophosphate 0 6 2 8 0 2 9
Organic ester:
4MU-Lignocerate 0 1 0 8 1 1 0
Abbreviation: Z, the N-terminal carbobenzoxy-group of
endopeptidase substrates.
*a= Corynebacterium variabilis e= G. terrae
b= Gordona bronchialis f= Tsukamurella paurometabola
c= G. sputi g= T. wratislaviensis
d= G. rubropertinca
--------------------------------------------------------------
Despite these improvements in classification, gordonae, rhodococci and tsukamurellae are difficult to identify though many tests have been recommended for this purpose. However, little confidence can be placed in the predictiveness of such tests due to the lack of congruence found between the results from different studies and the failure of investigators to examine representative strains of all validly described species.
The introduction of the genera Gordona(25) and Tsukamurella(26), for actinomycetes previously considered as rhodococci, leaves Rhodococcus as a homogeneous genus encompassing thirteen taxospecies most of which correspond with genomic species(27). It is most encouraging that the fluorogenic probes prepared from 4-methylumbelliferone and 7-amino-4-methylcoumarin have been shown to demonstrate a range of putatively diagnostic enzyme activities in suspensions of non-growing rhodococcal cultures (Table 1). Similar features can be provisionally weighted for the identification of Corynebacterium, Gordona and Tsukamurella species (Table 2). The species Coryne- bacterium variabilis now includes the organism previously known as Caseobacter polymorphus(28).
The ability to characterise and identify mycolic acid containing actinomycetes notably rhodococci, is growing given their increasing importance in agriculture, medicine and industry(24). Rhodococci, and possibly gordonae and tsukamurellae, are opportunistic pathogens of human and animals. These organisms produce bioactive compounds, effect useful xenobiotic transformations of complex molecules, and degrade diverse pollutants, including nitrate, phthalate esters and polychlorinated phenols. Consequently rhodococci may have a role to play in bioremediation and detoxification of contaminated soils.
In future, the availability of automated data acquisition systems will facilitate the detection of enzyme activities in bacteria, including actinomycetes. Microscan (Sacramento, California, U.S.A.) and Sensititre Ltd. (East Grinstead, West Sussex, U.K.) manufacture kits containing fluorogenic enzyme tests. The conjugated substrates are contained in microtitre trays and quantitative results of fluorescent intensity are obtained using a computer controlled fluorimetric plate reader. Raw data are interpreted using appropriate computer software. Conclusions The first step in the identification of actinomycetes is usually the assignment of organisms to genera and families. This can be achieved for most broad groups using a combination of chemotaxonomic and morphological properties. Similarly, fluorogenic probes based on 7-amino-4-methylcoumarin and 4- methylumbelliferone can provide valuable data for the identification of actinomycete species. An important objective in actinomycete systematics should be a thorough examination of representative strains from well circumscribed species to highlight diagnostic enzymes that can be used to identify organisms to the rank of species. References
1) Williams, S.T., J.C.Vickers & M.Goodfellow: Application of new theoretical concepts to the identification of streptomycetes. In "Computer-Assisted Bacterial Identification" (eds. M. Goodfellow, D. Jones and F.G. Priest), pp. 289-306, Academic Press, London, 1985.
2) O'Donnell, A.G.: Recognition of novel actinomycetes. In "Actinomycetes in Biotechnology" (eds. M. Goodfellow, S.T. Williams and M. Mordarski), pp. 69-88, Academic Press, London, 1988.
3) Goodfellow, M.: Suprageneric classification of actinomycetes. In "Bergey's Manual of Systematic Bacteriology. Volume 4" (eds. S.T. Williams, M.E. Sharpe and J.G. Holt), pp. 2333-2339, Williams & Wilkins, Baltimore, 1989.
4) Lechevalier, H.A.: A practical guide to generic identification of actinomycetes. In "Bergey's Manual of Systematic Bacteriology, Volume 4" (eds. S.T. Williams, M.E. Sharpe and J.G, Holt), pp. 2344-2347, Williams & Wilkins, Baltimore, 1989.
5) Goodfellow, M. & D.E.Minnikin (eds.): Chemical Methods in Bacterial Systematics. Academic Press, London, 1985.
6) O'Donnell, A.G., D.E.Minnikin & M. Goodfellow: Integrated lipid and wall analysis of actinomycetes. In "Chemical Methods in Bacterial Systematics" (eds. M. Goodfellow and D.E. Minnikin), pp. 131-143, Academic Press, London, 1985.
7) Schaal, K.P.: Identification of clinically significant actinomycetes and related bacteria using chemical techniques. In "Chemical Methods in Bacterial Systematics" (eds. M. Goodfellow and D.E. Minnikin), pp. 359-381, Academic Press, London, 1985.
8) Goodfellow, M. & A.G.O'Donnell: Search and discovery of industrially significant actinomycetes. In "Microbial Products: New Approaches" (eds. S. Baumberg, I.S. Hunter and P.M.Rhodes), pp. 343-383, Cambridge University Press, Cambridge, 1989.
9) Langham, C.D., S.T.Williams, P.H.A. Sneath & A.M.Mortimer: New probability matrices for identification of Streptomyces. J. gen. Microbiol., 135: 121-133.1989.
10) Kersters, K. & J.De Ley: Enzymatic tests with resting cells and cell free extracts. In "Methods in Microbiology, Volume 6A" (eds. J.R.Norris and D.W.Ribbons), pp. 44-65, Academic Press, London, 1971.
11) Kilian, M.: Rapid identification of Actinomycetaceae and related bacteria. J. Clin. Microbiol., 8: 127-133, 1978.
12) Schofield, G.M. & K.P.Schaal: Rapid micromethods for detecting deamination and decarboxylation of amino acids, indole production and reduction of nitrate and nitrite by facultatively anaerobic actinomycetes. Zbl. Bakt. Hyg., 1. Abt., Orig. A, 247: 383-391, 1980.
13) Goodfellow, M., E.G.Thomas, A.C.Ward & A.L.James: Classification and identification of rhodococci. Zbl. Bakt., 274: 299-315, 1990. 14) Goodfellow, M., J.Zakrzewska-Czerwinska, E.G.Thomas, M.Mordarski, A.C. Ward & A.L. James: Polyphasic taxonomic study of the genera Gordona and Tsukamurella including the description of Tsukamurella wratislavien- sis sp.nov. Zbl. Bakt., (in press).
15) Kilian, M. & P.Bulow: Rapid identification of Enterobacteriaceae. I. Detection of bacterial glycosidases. Acta Path. Microbiol. Scand., Sect. B, 84: 245-251, 1976.
16) Freney, J., M.T.Duperron, C.Courtier, W.Hansen, F.Allard, J.M.Boeufgras, D.Monget & J.Fleurette: Evaluation of ALI Coryne in comparison with conventional methods for identifying coryneform bacteria. J. Clin. Microbiol., 29: 38-41, 1991.
17) Grange, J.M.: Fluorometric assay of mycobacterial group specific hydrolase enzymes. J. Clin. Pathol., 31: 378-381, 1978
18) Grange, J.M. & K.Clark: Use of methylumbelliferone derivatives in the study of enzyme activities of mycobacteria. J. Clin. Pathol., 30: 151-153, 1977.
]9) Goodfellow, M., T.M.Embley & B. Austin: Numerical taxonomy and emended description of Renibacterium salmoninarum. J. gen. Microbiol., 131: 2739-2752, 1985.
20) Goodfellow, M., C.Londsale, A.L.James & O.C.MacNamara: Rapid biochemical tests for the characterisation of streptomycetes. FEMS Microbiol. Lett., 43: 39-44, 1987.
21) Goodfellow, M., E.G.Thomas & A.L. James: Characterisation of rhodococci using peptide hydrolase substrates based on 7-amino-4-methylcoumarin. FEMS Microbiol. Lett., 44: 349-355, 1987.
22) Slifkin, M. & G.M.Gill, Rapid biochemical tests for the identification of groups A, B, C, F and G streptococci from throat cultures. J. Clin. Microbiol., 18: 29-32, 1983.
23) Goodfellow, M. & T.Cross: Classification. In "The Biology of the Actinomycetes" (eds. M Goodfellow, M. Mordarski and S.T. Williams), pp. 7-164, Academic Press, London, 1984.
24) Goodfellow, M.: The family Nocardiaceae. In "The Prokaryotes, Second Edition" (eds. A. Balows, H.G.Truper, M.Dworkin, W.Harder and K.H.Schleifer. Springer Verlag, New York, (in press).
25) Stackebrandt, E., J.Smida & M.D. Collins: Evidence of phylogenetic heterogenicity within the genus Rhodococcus.: Renewal of the genus Gordona (Tsukamura). J. Gen.Appl.Microbiol., 34:34 1-348, 1988.
26) Collins, M.D., J.Smida, M.Dorsch & E. Stackebrandt: Tsukamurella gert.nov. harboring Corynebacterium paurometabolum and Rhodococcus aurantiacus. Int. J. System. Bact., 88: 385-391, 1988.
27) Zakrzewska-Czerwinska, J., M.Mordarski & M.Goodfellow: DNA base composition and homology values in the classification of some Rhodococcus species. J. gen. Microbiol., 134: 2807-2813, 1988.
28) Collins, M.D., J.Smida & E.Stackebrandt: Phylogenetic evidence for the transfer of Caseobacter polymorphus (Crombach) to the genus Corynebacterium. Int. J. System.Bact., 19: 7-9, 1989. Copyright 1996 C.E.T.A., The International Centre for Theoretical and Applied Ecology, Gorizia The following images related to this document are available:Line drawing images[ac91008b.gif] [ac91008a.gif] |
| |||||||||
{kind=link}
{kind=link}