 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Towards a molecular identification of Streptomyces species in pure culture and in environmental samples. ERKO STACKEBRANDT(l), WERNER LIESACK(l), RICK WEBB(1,2) and DAGMAR WITT(3) (l)Department of Microbiology, University of Queensland, St. Lucia, Queensland 4072, Australia; (2)Electron Microscope Centre, University of Queensland, St. Lucia, Queensland 4072, Australia; (3)Institut fur Allgemeine Mikrobiologie, Christian-Universitat, 2300 Kiel, Federal Republic of Germany
Code Number: AC91012
Size of Files:
Text: 26.8K
Graphics: line drawings (gif) - 12.9K
photographs (jpg) - 221.4K
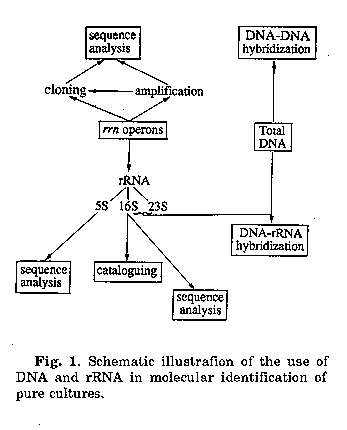
During the last 5 years a number of scientific reports have been published that demonstrate the value of molecular techniques in the circumscription of microbial populations in the environment. The needs for genetic characterization has been called for when it became obvious that traditional phenotype based techniques failed to detect the full diversity of bacterial types and to isolate the majority of organisms present in a given sample, e.g. it has been speculated that the percentage of organisms discovered in the environment does not exceed 20%(1). Besides the application of DNA hybridization, most modern studies are based on information of rRNA and rRNA genes, molecules that have intensively been dealt with during the last two decades. The approaches used today in several laboratories worldwide are summarized in Fig. 1.
Information obtained from comparative sequencing studies has been used to design and develop taxon-specific (mainly species-specific) oligonucleotide probes. They facilitate the identification of pure cultures and clones of gene libraries by Southern blot hybridization and they are able to visualize individual target cells within a mixed population by epifluorescence microscopy and eventually by flow cytometry. Only recently, sequence information has also been used to develop primers to be applied in the polymerase chain reaction (PCR)mediated amplification of rrn genes which allows to copy the sequence enclosed between the two PCR primers up to million fold.
Here we summarize the information that is available for the molecular identification of streptomycetes. These studies have been initiated to learn more about the role these organisms play in the food web, to monitor naturally occurring, and eventually genetically engineered released strains in the environment and to identify new antibiotic producing strains. The genera Streptomyces and Streptoverticillium have recently been unified in Streptomyces(2). Consequently, the new definition of Streptomyces will be used in the following text.
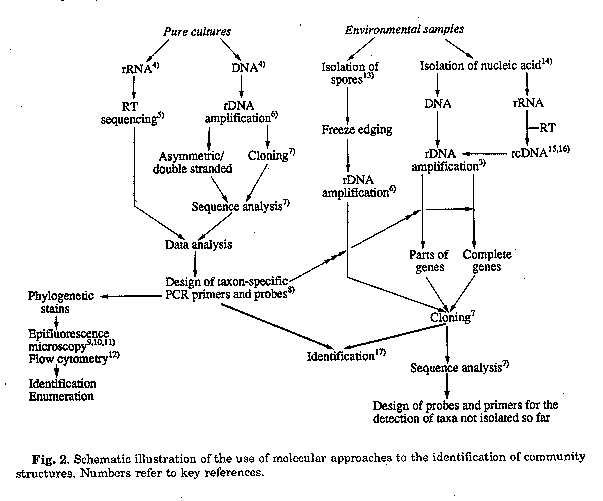
It is beyond the scope of this communication to give detailed protocols of all the procedures mentioned. The latest applications have just recently be comprehensively compiled in the series "Modern Microbiological Methods"(3). The main strategies are rather outlined in a flow diagram which also indicates the main sources of techniques and contributions to the molecular identification of bacteria. Basically two main streams emerge of which the studies of pure cultures are prerequisite for a successful continuation of stage 2, viz. the monitoring of environmental samples.
Step 1. It is the goal to identify taxon specific stretches that can be used as targets for either oligonucleotide probes or primers. Although the comparison of the target stretch with a large data base may yield information' about the uniqueness of a sequence and conse- quently about the chance to hybridize specifically, a thorough testing of the probes on a large variety of highly to moderately related species is absolutely necessary. Considering certain parameters, one or two nucleotides differences between two target regions of 20 nucleotides in length are usually sufficient to obtain a discriminating signal between the homologous and the heterologous target in dot blot hybridization. On the other hand, since oligonucleotide primers used in PCR assays can not be reassociated to the targets at the same stringency than probes, primers need to be more selective.
A large selection of different labeling procedures have been described, and the non-radioactive, fluorescent dyes are slowly replacing radioactive label. Because of the universal application of probes and primers, some of these oligonucleotides are already commercially available and culture collections are going to establish probe banks for identification purposes.
The main problem encountered in step 2 is the isolation of high-molecular weight DNA directly from an environmental sample. Isolation of nucleic acids recovered from cells that are collected from the sample may eventually be easier but includes the danger of loosing information of those cells that can not be recovered under the methods used (up to 80%). Once DNA has been obtained, all subsequent steps, including PCR, cloning, detection of positive recombinants and sequence analysis follow described methods but cannot as yet be considered routine at all. Isolation of rRNA from environmental samples appears to be easier but rRNA has to be circumscribed into rcDNA prior to further manipulations. Due to the intrinsic property of the reverse transcriptase, however, certain post-transcriptional modifications causes the enzyme to leave the rRNA target. This results in the formation of a rcDNA fragment whose length is determined by the position of the primer and its nearest 5' modified nucleotide. Information about the presence and the location of these modifications in members of the group under investigation is therefore necessary to meet this problem with a different strategy. Gene libraries can be hybridized with probes of different taxon specificity which could eventually even lead to a semi-enumeration of species, e.g. by subsequent use of an universal probe to check positive, a 16S rrn gene(or part thereof) containing clone, a genus-specific probe to determine the fraction of its members within the total population, and a species specific probe to determine the fraction of this species within the total number of members of the genus. Clones that hybridize with a genus-specific probe but fail to hybridize with the whole range of available species-specific probes can be sequenced for identification purposes. Information from uniques stretches can be used to develop probes that in turn may allow the identification of the species grown in enrichment cultures of the respective environmental sample. The potential of the 5S rRNA Two 5S rRNA sequences of streptomycetes have been published so far(18,19) which differ from each other in only two base exchanges. It is therefore as yet impossible to determine whether or not those regions, known to contain variable stretches in prokaryotic 5S rRNAs are sufficiently different among streptomycetes to serve as species-specific target regions. However, considering the overall high degree of relationship found among streptomycetes, the conserved character of the 5S rRNA and its small size, it appears unlikely that the small rRNA species contains sufficient variations to serve as targets for species-specific probes. The potential of the 16S rRNA/rDNA Full sequences of 3 species and almost complete sequences of additional 14 species of Streptomyces have been published(20). Since the species investigated are representatives of several of the major clusters defined by numerical analyses(21), it is assumed that a broad spectrum of different genotypes had been included. The relative phylogenetic closeness of all actinomycetes makes it difficult to define a genus specific probe. The only discriminating position found in the 16S rRNA which clearly separate streptomycetes from all other actinomycetes is nucleotide 929 (C-residue). A probe developed to target a stretch around position 929 was highly discriminating at hybridization temperatures near the Tm value of the homologous DNA/rRNA duplex. The probe was able to detect all Streptomyces (incl. Streptoverticillium, the former genera Chainia and Kitasatoa, as well as Kitasatosporia). This probe will be of great value in rapid screening of gene libraries for streptomycetes 16S rDNA genes(l7).
Basically three regions could be detected which showed a higher degree of variability than the other parts of the molecules in which differences where more dispersed. These regions were named alpha (982 through 998), beta (1102 through 1122) and gamma (170 through 200). Analysis of the alpha and beta region of additional 60 strains from 40 Streptomyces species indicated an even higher degree of variation. Nevertheless, these regions were not sufficiently different to distinguish each of the species. 23 species of the former genus Streptoverticillium exhibited the same alpha/beta combination, 16 species and several isolates from the marine environment were characterized by a unique combination while nine other combinations were found in clusters of up to seven strains. Consequently, only 23 strains of the 77 strains investigated could be detected by a single probe or by a combination of two probes targeting these regions. For the design of PCR primers however these regions may be of significance. Their position in the 3' half of the molecule will allow a selective amplification of rDNAs of members from defined groups of streptomycetes (depending upon the primer used) when the assay contains a reverse primer that is located at the 5' terminus of the molecule (i.e. around position 15). Even more selective, eventually species - spe- cific PCR products will be obtained when the reverse primer targets the gamma region. This stretch reveals the highest degree of sequence variation. Of the 17 strains sequenced, 16 showed differences of up to 15 nucleotides. This stretch may carry the greatest potential of the 16S rRNA for the design of species- and eventually strain-specific probes to be used directly for screening colonies, gene libraries or those PCR products which enclose this target region. The potential of the intracistronic rrn spacer Sequences of the intracistronic spacer of only three Streptomyces species are as yet available, namely S. lividans(22), S. coelicolor A3(2)(23), and S. ambofaciens(19), The spacer range between 270 and 300 nucleotides in length. A partial sequence of about 100 nucleotides from the 3' terminus is also available from S. albus (Liesack, unpublished), one of the most distantly related species within the 16S rRNA tree(2) while at the 16S rRNA level S.lividans and the A3(2) strain are almost indistinguishable, the two strains of these species show significant differences within the primary structure of the spacer (about 12%) which are mainly located in two regions. 74 through 99 and 119 through 144. Even more dramatic is the difference between these strains and S. ambofaciens (about 30%). Interestingly, the 40 or so nucleotides of the 3' terminus are absolutely identical for all of the four species investigated (but different to the homologous region in Mycobacterium leprae). It appears that the less conserved spacer carries a high potential to contain information for both species- and genus-specific probes and primers. Sequence analysis of fragments, obtained after amplification of this region by PCR, using the reverse primer and the RT primer directed against the highly conserved 3' termini of the 16S rDNA and the spacer, respectively, should give rapid information about the extent to which the individual species, or eventually even strains, differ from each other. Nothing is known as yet about the degree of intracistronic sequence variation of the spacer region within the genome of a strain. The potential of the 23S rRNA/rDNA Information about a rather variable stretch within the 23S rRNA is restricted to helix 54, that occurs in an extended version in all members of the order Actinomycetales, including six streptomycetes investigated so far(17). Up to 33 differences were found in members that could not be distinguished in the primary structure of the alpha region of their 16S rRNA. This region appears to be valuable for inter- nal species-specific probe target sides and for primers to be used either for PCR or sequence analysis of the 5' half (RT primer) and the 3' half (reverse primer) of the 23S rDNA. As already mentioned for the spacer, the full potential of this and similar variable regions of the 23S rRNA/DNA can only be evaluated after much more extensive and comparative sequence analyses. Amplification of rDNA from Streptomyces albus spores
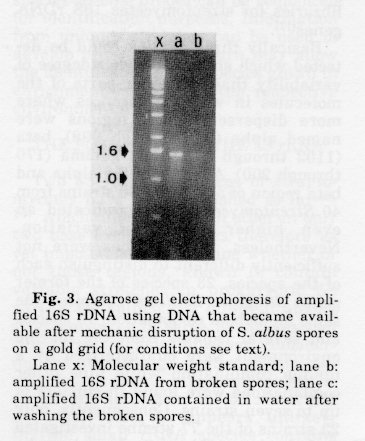
Spores of S. albus were isolated from an agar culture by filtration through a cotton wool pad located in the tip of a 10 ml syringe, washed three times with sterile water and stored at -20 C. Specimens were fixed in 2% paraformaldehyde, 0.5% glutaraldehyde in 0.1M cacodylate buffer. Following a washing step in the same buffer they were embedded in 10% gelatin in phosphate buffer. Free aldehydes were quenched in 50 mM glycine in phosphate-buffered saline for 150 min. Specimen were infiltrated with 2.3 M sucrose and frozen by plunging in liquid nitrogen. Sections (ca. 150 nm thick) were cut on a Reicher-Jung FC4 cryo-ultra-microtome, placed on Formvar coated gold grids and stored on 2% gelatine, 0.25% agarose at 4 C. Prior to 16S rDNA gene amplification a grid, containing about 104 spores, was washed in 700 ul sterile water and a 50 ul sample thereof tested for the presence of DNA that has been released during the washing procedure. The grid was submerged into 100 ul PCR reagent buffer in an 0.5 ml vial and the experiment done as de- scribed(24), using the universal primers 9-27 and 1525-1544. Fig. 3 clearly shows a strong band of amplified S. albus spore DNA and a weaker band of amplified DNA released into the water during the washing step. The control for contami- nation (same PCR assay but omitting DNA) was negative which indicates that the DNA fragments obtained actually originated from the broken spores.
Lane x: Molecular weight standard; lane b: amplified 16S rDNA from broken spores; lane c: amplified 16S rDNA contained in water after washing the broken spores.
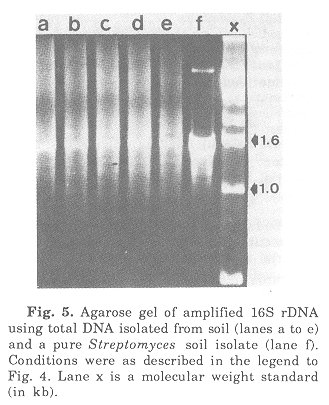
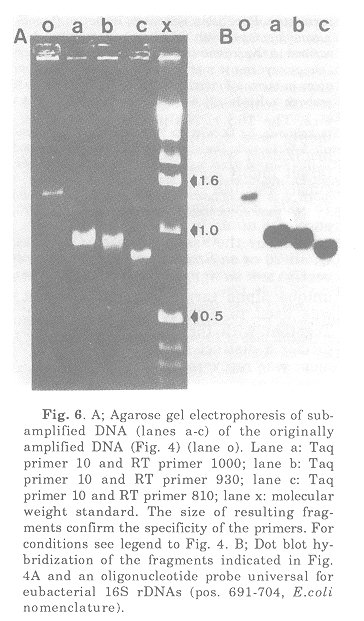
In order to test the specificity of PCR primers to be used in the amplification of almost complete 16S rRNA genes they were tested against pure DNA from Xenorhabdus luminescens, Bacillus subtilis and Streptomyces scabies. 16S rDNA was amplified using primers targeting positions 9-27 and 1525-1544. Conditions for annealing and amplification have been reported(23). The fragments were monitored by agarose gel electrophoresis (Fig. 4). As compared to the molecular weight standard the amplification products had the expected size of about 1.5 kb. The following is a brief summary of preliminary results of amplification studies on DNA isolated from soil. DNA isolation followed published procedures(14). The first step included an enrichment of those eubacterial 16S rRNA genes which could be amplified with the primers used (Fig. 5).
Fig. 5. Agarose gel of amplified 16S rDNA using total DNA isolated from soil (lanes a to e) and a pure Streptomyces soil isolate (lane Conditions were as described in the legend to Fig. 4. Lane x is a molecular weight standard (in kb).
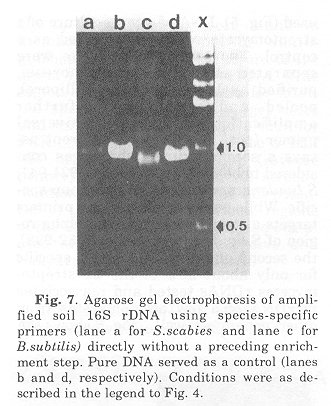
Fig. 7. Agarose gel electrophoresis of amplified soil 16S rDNA using species-specific primers (lane a for S.scabies and lane c for B.subtilis) directly without a preceding enrichment step. Pure DNA served as a control (lanes b and d, respectively). Conditions were as described in the legend to Fig. 4.
Conclusion The results not only point towards the existence of streptomycetes in the soil sample investigated but also indicate that the use of more specific primers will enable us to identify groups of species, if not species. At the time of these studies information about the gamma region of the 16S rRNA, the intracistronic spacer and the 23S rRNA were not available. The respective probes and primers are currently under investigation. Application of these molecular techniques, together with the possibility to amplify rDNA from thin sections of spores and mycelium, and in combination with classical enrichment studies, will in the long run signifi- cantly increase our understanding about the role streptomycetes play in the environment. Acknowledgements. We would like to thank the organizers of the IUMS satellite meeting "New Impact of Actinomycetes Taxonomy", Takeda Chemicals Ind. Ltd., Osaka, September 14, 1990 to give one of us (E.S.) the opportunity to present data on the development of gene probes. Parts of this research was supported by a grant from the Australian Research Council. References 1) Woese, C.R.: Bacterial evolution. Microbiol. Rev., 51: 221-271, 1987.
2) Witt, D. & E.Stackebrandt: Unification of the genera Streptoverticillium and Streptomyces, and amendation of Streptomyces Waksman and Henrici 1943, 339AL. System. Appl. Microbiol., 13: 361-371,1990.
3) Stackebrandt, E. & M.Goodfellow (eds.): Nucleic Acid Techniques in Bacterial Systematics. John Wiley & Sons, Chichester, 1991.
4) Johnson, J.L.: Isolation and purification of nucleic acids. In "Nucleic Acid Techniques in Bacterial Systematics" (eds. E. Stackebrandt & M. Goodfellow), pp. 1-20, John Wiley & Sons, Chichester, 1991.
5) Lane, D.J.: 16S/23S rRNA sequencing. In "Nucleic Acid Techniques in Bacterial Systematics" (eds. E.Stackebrandt & M. Goodfellow), pp. 115-176, John Wiley & Sons, Chichester, 1991.
6) Giovannoni, S.: The polymerase chain reaction. In "Nucleic Acid Techniques in Bacterial Systematics" (eds. E.Stackebrandt & M.Goodfellow), pp. 177-204, John Wiley & Sons, Chichester, 1991.
7) Ludwig, W.: DNA sequencing in bacterial systematics. In "Nucleic Acid Techniques in Bacterial Systematics" (eds. E.Stackebrandt & M.Goodfellow), pp. 69-94, John Wiley & Sons, Chichester, 1991.
8) Stahl, D.A. & R.Amann: Development and application of nucleic acid probes. In "Nucleic Acid Techniques in Bacterial Systematics" (eds. E.Stackebrandt & M.Goodfellow), pp. 205-248, John Wiley & Sons, Chichester, 1991.
9) DeLong, E.F., S.Wickham & N.R.Pace: Ribosomal RNA-based probes for the identification of single cells. Science, 243: 1360-1363, 1989.
10) Giovannoni, S.J., T.B.Britschgi, C. Moyer & K.G.Field: Genetic diversity in Sargasso Sea bacterioplankton. Nature, 345: 6062, 1990.
11) Amann, R.I., L.Krumholz & D.A.Stahl: Fluorescent oligonucleotide probing of whole cells for determinative, phylogenetic and environmental studies in microbiology. J. Bact., 172: 762-770, 1990.
12) Amann, R.I., B.J.Binder, R.J.Olson, S.W.Chisholm, R.Devereux & D.A.Stahl: Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations. Appl. Environ. Microbiol., 56: 1919-1925, 1990.
13) Herron, P.R. & E.M.H.Wellington: New method for extraction of streptomycete spores 'from soil and application to the study of lysogeny in sterile amended and nonsterile soil. Appl. Environ. Microbiol., 56: 1406-1412, 1990.
14) Ogram, A., G.S.Sayler & T.Barkay: The extraction and purification of microbial DNA from sediments. J. Micro. Meth., 7: 57-66, 1987. 15) Ward, D.M., R.Weller & M.Bateson: 16S rRNA sequences reveal numerous uncultured microorganisms in a natural community. Nature, 345: 63-65, 1990.
16) Kemmerling, C., C.Witt, W.Liesack, H.Weyland & E.Stackebrandt: Approaches for the molecular identification of streptomycetes in marine environment. In "Current Topics in Marine Biotechnology" (eds. S.Miyachi, I.Karube & Y.Ishida), pp. 423-426, Fuji Technology Press Ltd., Tokyo, 1989.
17) Stackebrandt, E., D.Witt, C.Kemmerling, R.Kroppenstedt & W.Liesack: Designation of streptomycete 16S and 23S rRNA-based target regions for oligonucleotide probes. Appl. Environ. Microbiol., (in press).
18) Wolters, J. & V.A.Erdmann: Compilation of SS rRNA and SS rRNA gene sequences. Nucl. Acids Res., 16: rl-r70, 1988.
19) Pemodet, J.L., M.T.Alegre, F.Boccard & M.Guerineau: Organization and nucleotide sequence analysis of a ribosomal RNA gene cluster from Streptomyces ambofaciens. Gene, 79: 3346, 1989.
20) Williams, S.T., M.Goodfellow, G.Alderson, E.M.H.Wellington, P.H.A.Sneath & M.J. Sackin: Numerical classification of. Streptomyces and related genera. J. gen. Microbiol., 129: 1743-1813, 1983.
21) Suzuki, Y., Y.Ono, A.Nagata & T. Yamada: Molecular cloning and characterization of an rRNA operon in Streptomyces lividans TK21. J. Bacteriol., 170: 1631-1636, 1988.
22) Baylis, H.A. & M.J.Bibb: Transcriptional analysis of the 16S rRNA gene of the rrnD gene set of Streptomyces coelicolor A3(2). Mol. Microbiol., 2: 569-579, 1988.
23) Liesack, W., M.A.O.H. Menke & E. Stackebrandt: Rapid generation of vector-free Digoxygenin-dUTP labelled probes for nonradioactive hybridization using the polymerase chain reaction (PCR) method. Syst. Appl. Microbiol., 13: 255-256, 1990. Copyright 1996 C.E.T.A., The International Centre for Theoretical and Applied Ecology, Gorizia
The following images related to this document are available:Photo images[ac91012g.jpg] [ac91012d.jpg] [ac91012e.jpg] [ac91012c.jpg] [ac91012f.jpg]Line drawing images[ac91012b.gif] [ac91012a.gif] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}