 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
CLONING OF SINEFUNGIN BIOSYNTHETIC GENES FROM STREPTOMYCES INCARNATUS TO STREPTOMYCES LIVIDANS HALINA MALINA(1), FRANCOISE LAWRENCE, and MALKA ROBERT-GERO Institut de Chimie des Substances Naturelles, CNRS, 91198 Gif-sur-Yvette Cedex, France. (1) Present address: Department of Ophthalmology, University of Zurich, R/irnistrasse 100, CH-8091 Zurich, Switzerland
Code Number: AC95006
Sizes of Files:
Text: 20K
Graphics: Line Drawings (gif) 31K
Photos (jpg) 21K / Halftones (gif) 33K
ABSTRACT. Biosynthetic genes (sin genes) of
sinefungin, a transmethylase inhibitor, antifungal and
antiparasitic nucleoside antibiotic produced by
Streptomyces incarnatus, were shotgun cloned into
Streptomyces lividans. The genes are clustered on a 14
kb fragment of DNA. The expression of the sin gene
cluster in S. lividans leads to the production of a
sinefungin-like antibiotic. The compound has antifungal and
antiparasitic activities in vitro. The expression of
the gene cluster in S.lividans and E. coli
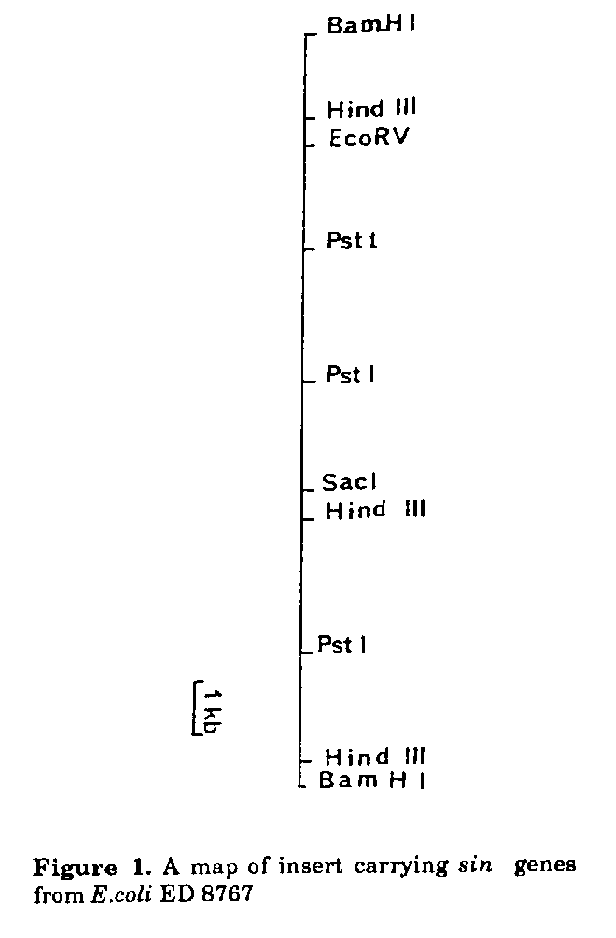
causes an astringent-like response.Sinefungin, a nucleoside antibiotic characterized by a relatively simple biosynthetic pathway (Malina et al., 1987; Parry et al., 1989), inhibits trans- methylation in vitro and in vivo (Blanchard et al., 1986). Inhibition of DNA methylation by sinefungin in vitro has been observed for Streptomyces antibioticus but not for S.incarnatus, the producing strain (Barbes et al., 1990). Cloning of antibiotic biosynthetic genes and studying their expression in heterologous hosts provide a tool to ob- tain modified antibiotics (Huchinson et al., 1989). Since 1984, the genes of several antibiotic biosynthetic pathways have been cloned (Malpartida and Hopwood, 1984), and from these data it appears that they are clustered with the antibiotic-resistance genes (Chater, 1990). Here we describe the interspecific cloning of the sinefungin biosynthetic pathway from S.incarnatus into S. lividans. MATERIALS AND METHODS Bacterial strains and plasmids. S. lividans, TK-24 str-6, was used as a Streptomyces-cloning vector host. E. coli, ED8767 (met B, rec A, hsd S), (Murray et al., 1977) was used as host for recombinant E.coli-Streptomyces plasmids, and E. coli plasmid pBR322 for the construction of the recombinant plasmids E. coli- Streptomyces. Saccharomyces pastorianus and Leishmania donovani promastigotes were used to test the antibiotic activity of the strain after cloning. Culture conditions and transformations procedures. Streptomyces protoplast preparation, transformation and regeneration were performed as described by Hopwood et al. (1985). Media A, used for the control of sinefungin expression, and C, for the of antibiotic activity, were previously de- scribed (Malina et al., 1985). Antibioticresistant transformants were selected on NE medium (Murakami et al., 1986) containing neomycin (Aph) (1 ug/ml) and thiostrepton (Tsr) (50 ug/ml). The E.coli strain was grown in LB medium (Maniatis et al., 1982). The transformants were selected on LB agar containing tetracycline (Tc) (12.5 ug/ml) and ampicillin (Ap) (100 ug/m|). DNA isolation and manipulations. Total Streptomyces DNA was extracted as described by Hunter (1985). The Streptomyces plasmid vector pIJ61 was isolated by.alkaline lysis (Kieser, 1984) and the plasmid containing cloned sin genes, according to the method of Murakami et al. (1983) followed by centrifugation in CsCl. E.coli plasmids were isolated as described by Maniatis et al. (1982). Dephosphorylation and ligation of the plasmid DNA were performed according to Hopwood et al. (1985). Restriction enzymes were used manufacturers' (Pharmacia) instructions. Agarose electrophoresis was performed in TAE buffer in 0.8% agarose (Maniatis et al., 1982). DNA-DNA hybridization. DNA fragments separated on agarose gel were transferred to Hybond N membranes (Amersham) in the presence of 20 x SSC. Membranes were dried at room temperature and DNA was crosslinked by UV exposure (3 min at 312 nm). Prehybridisation was carried out at 42 C in a buffer containing 50% formamide, 0.1% SDS, 2 x Denhardt's solution, 5 x SSPE (Maniatis et al., 1982) and denatured calf thymus DNA (100 ug/ml). The probe was labelled by random priming (Feinberg and Vogelstein, 1983). After 2 hrs the membranes were placed in the same buffer containing the alpha-^32P-labelled denatured DNA probe (13x10^6 cpm/ml) and hybridisation was carried out for 24-48 hrs at 42 C. Membranes were washed sequentially, twice in 2 x SSPE 0.1% SDS (w/v) at room temperature for 10 min, then in 0.1% SSPE (w/v) at 50 C for 15 min and finally in 0.1 x SSPE 0.1% SDS (w/v) at 50 C for 10 min. Autoradiography was carried out by exposure to Hyperfilm MP (Amersham) at -70 C for 4-7 dd. Isolation and analysis of the antibiotic after cloning. The antibiotic was isolated according to the method used for sinefungin isolation (Rhone Poulenc Patent, France 76111141, 15.04.1976) with some modifications. The fermentation broth was acidified to pH 3.5 and centrifuged. The antibiotic was isolated in succession from the supernatant on Amberlite IR120, Amberlite XAD-2, Amberlite IRC50 and by silica gel chromatography. The activity was tested at all steps of purification by TLC bioautography against Saccharomyces. pastorianius. TLC was performed on silicagel plates with methanol: chloroform : 28% NH4OH (3:1:1 v/v). Compounds were visualised by UV light, ninhydrin and bioautography. HPLC analysis was performed as previously described (Malina and Gero, 1988) using 0.05 mM ammonium acetate as mobile phase. Analysis of the purified antibiotic was performed by UV, IR, NMR 400 MHz, and FAB. Protein analysis. The mycelium from a 72 hr culture in A medium was labelled with 35S-methionine (10 uCi/ml) for 1 hr. Cells were harvested by centrifugation, washed twice in phosphate buffer (pH 7) and subjected to ultrasounds at 4 C. Cells were further lyzed by addition of 1 volume of 2 x solu- bilisation buffer containing 0.2 M Tris HCl pH 7, 0.4% SDS, 20% (v/v) glycerol and 2mM phenylmethylsulfonyl fluoride, and heated in a boiling-water bath for 3 min. Samples were electrophoresed in 10% acrylamide gels (Laemmli, 1970). Gels were stained with Coomassie blue, dried under vacuum and autoradiographed on Amersham MP films for 8-15 dd. Molecular weight of the polypeptides was estimated by coelectrophoresis with standard proteins. Methionine uptake and incorporation into proteins were obtained from the radioactivity of cold TCA soluble and insoluble materials respectively. Protein concentration was determined according to Bradford (1976) procedure, using bovine serum albumin as standard. RESULTS AND DISCUSSION S.incarnatus harbours 5 low copy plasmids of different sizes, named pGIF 1-5 (Malina and Gero, 1990; 1992). Transformations experiments showed that the structural biosynthetic genes are not plasmid encoded. Therefore, the cloning of sin biosynthetic genes was carried out from S.incarnatus chromosomic DNA into S. lividans. Generally for cloning biosynthetic pathways it is easier to start with the resistance genes, but in the present case this was not possible due to the lack of sensitivity of the prokaryotes (Streptomyces spp., and E.coli) to extracellular sinefungin. S. incarnatus DNA was partially digested by Mbol and ligated with a pIJ61 vector (Aph^RTsr^R) (Thompson et al., 1982) which had been previously cleaved with BamHl and dephosphorylated. The resulting DNA was introduced into S. lividans protoplasts. Transformants with phenotype Aph^S Tsr^R were screened for production of antifungal metabolites. Two out of 2300 colonies examined were found to inhibit the growth of Saccharomyces pastorianus. Cultivation of these transformants, named S. lividans LI-41 and S. lividans LI-42, was difficult to carry out due to the rapid protoplast formation from the mycelium. The plasmid copy number was significantly reduced in comparison to the pIJ61, probably due to deletion or loss of the plasmids during the protoplast formation. The fermentation broth of the variants showed antifungal activity against Saccharomyces pastorianus and Candida albicans and antiparasitic activity against Leishmania donovani promastigotes in vitro. HPLC analysis of the active post-fermentation broth showed the presence of new substances, as compared to that produced by the host strain containing the cloning vector pIJ61. Only one substance (PM-4), having a retention time of 3.69 min on HPLC, similar to sinefungin (3.76 min) however was active. The compound has maximum absorbance at 257 nm and is ninhydrin positive. The 1H-NMR 400 MHz spectrum showed the presence of the 2H and 8H protons of adenine and the protons of ribose and of the side chain. The molecular mass of the compound however was 364, whereas that of sinefungin is 381. According to these results the expression of the sin gene cluster in S.lividans appears to lead to the production of a modified nucleoside antibiotic. The exact structure of this substance could not be evaluated due to instability of the plasmid and thus to loss of strain activity. When the plasmid was isolated from S. lividans LI-41, only small quantities of plasmid DNA (100 ng per 200 ml cul- ture) were obtained. All efforts to isolate the plasmids by alkaline lysis, suitable for the plasmid vector pIJ61, failed. The analysis of the BamHl digest of the plasmid DNA on agarose gels showed the presence of a plasmid vector of 15 kb and numerous inserts having sizes 2 kb to 24 kb. The plasmid DNA from S. lividans LI-41, digested by BamHI and labelled by random priming, hybridised strongly with the S. incarnatus DNA but not with heterologous DNA (Fig. 1). Thus, the cloned fragment originated from S.incarnatus. However, the great fragmentation of the plasmid obtained from S.lividans LI-41 did not allow the construction of a map of insert. It might be that the instability of the plasmid associated to the sin gene expression results from altered host methylation (Vedel et al., 1978; Barras and Marius, 1989).
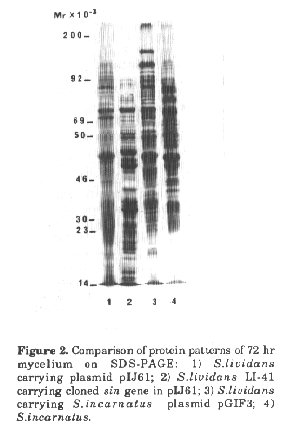
Figure 2. Comparison of protein patterns of 72 hr mycelium on SDS-PAGE: 1) S.lividans carrying plasmid pIJ61; 2) S.lividans LI-41 carrying cloned sin gene in pIJ61; 3) S.lividans carrying S.incarnatus plasmid pGIF3; 4) S.incarnatus.
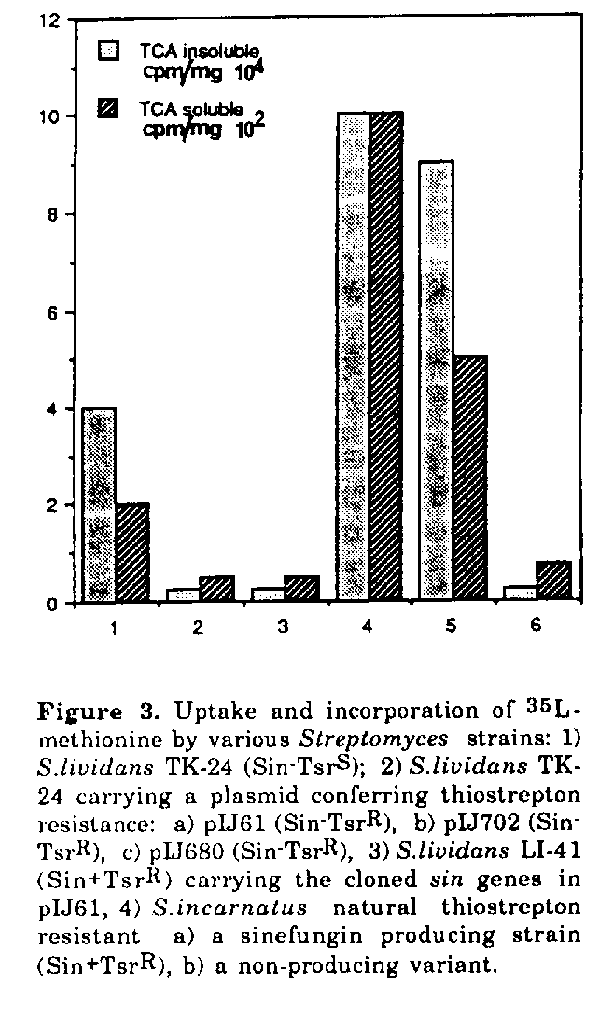
Expression of the sin gene cluster in S.lividans even on the low copy plasmid pIJ61 caused a strong stress response. Protoplast formation during growth was observed and the plasmid copy number was reduced strongly compared with plasmid pIJ61. The insert carrying the sinefungin gene cluster was progressively deleted. In comparison to the host strain the protein pattern was changed considerably. Some strongly expressed proteins (such as that of 47 kb) were not observed in the parental strain nor in S.lividans which harbours the plasmid of S.incarnatus and which does not produce active metabolites (Fig.2). Interestingly 35S-methionine uptake and incorporation was increased 10-20 fold in sinefungin producing strains (S.lividans LI-41 containing the cloned sin genes, and S.incarnatus) in comparison to non-producing ones (Fig.3).
However, the application of cloning to production of this kind of antibiotic is very difficult due to great plasmid instability. This suggests that cloning of molecules involved in regulation of key metabolites has to be carried out to- gether with the cloning of certain specific host factors. ACKNOWLEDGEMENTS. The authors thank Prof. N.E.Murray (University of Edinburgh) for the strain E.coli ED 8767 and Dr.Ph.Hendrickson for proof-reading the manuscript. The shotgun cloning of the sin biosynthetic genes was performed by H.M. in the laboratory of Prof. J.E.Davies, Department of Biotechnology, Pasteur Institute, Paris, to whom the senior author is grateful for advice and helpful discussions. This work was partially supported by a grant of the Medical Research Foundation, Paris. REFERENCES Barbes, C., J.Sanchez, M.Y.Yebra, M.RobertGero & C.Hardisson (1990). Effects of sinefungin and S-adenosylhomocysteine on DNA and protein-methyltransferases from Streptomyces and other bacteria. FEMS Microbiol. Lett., 69: 239-244 Barras, F. & M.G.Marinus (1989). The great GATC DNA methylation in E. coli. TIG, 5: 139-143 Biro, S. & K.F. Chater (1987). Cloning of Streptomyces griseus and Streptomyces lividans genes for glycerol dissimilation. Gene, 56:79-86 Blanchard, P., N.Dodic, J.LFourrey, M. Geze, F.Lawrence, H.Malina, P.Paolantonacci, M. Vedel, C.Tempete, M. Robert-Gero & E.Lederer (1986). Sinefungin and derivatives: synthesis, biosynthesis and molecular target studies in Leishmania. In: R.T.Borchardt, C.C.Creveling & P.M.Ueland (eds.) The Biochemistry of S-Adenosyl Methionine as a Basis for Drug Design. Humana Press, Inc., pp. 435446 Bradford, M. (1976). A rapid and sensitive method for the quantification of microgramm quantities of protein utilizing the principle of protein dye binding. Anal. Biochem., 72:248-254 Chater, K.F. (1990). The improving prospects for yield increase by genetic engineering in antibiotic producing Streptomyces. Biotechnology, 8: 115-121. Feinberg, A.P. & B.Vogelstein (1983). A technique for DNA restriction endonuclease fragments to high specific activity. Anal. Biochem., 132:6-9 Hopwood, D.A., M.J.Bibb, K.F.Chater, T. Kieser, C.J.Bruton, H.M.Kieser, D.J.Lydiate, C.P.Smith, J.M.Ward & H.Schrempf (1985). Genetic Manipulation of Streptomyces. A Laboratory Manual. The John Innes Foundation, Norwich Hunter, J.S. (1985). Gene cloning in Streptomyces. In: D.M.Glover (ed.) DNA Cloning. Vol. 11. A Practical Approach. IRL Press, Oxford, Washington, DC, pp. 19-44 Kieser, T. (1984). Factors affecting the isolation of CCC DNA from Streptomyces lividans and Escherichia coli. Plasmid, 12:19-36 Hutchinson, R.C., C.N. Borell, S.L.Otten, K.J. Stutzman-Engwall & Y.Wang (1989). Drug discovery and development the genetic engineering of antibiotic producing microorganisms. J. Med. Chem., 32:929-937 Kieser, T. (1984). Factors affecting the isolation of CCC DNA from Streptomyces lividans and Escherichia coli. Plasmid, 12:19-36 Laemmli, U.K. (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227:680-685 Malina H., C.Tempete & M.Robert-Gero (1985). Enhanced sinefungin production by medium improvement, mutagenesis and protoplast regeneration of Streptomyces incarnatus. J. Antibiot., 38:1204-1210 Malina, H., C.Tempete & M.Robert-Gero (1987). Biosynthesis of sinefungin by cell free extract. J. Antibiot., 40:505-511 Malina, H. & M.Robert-Gero (1988). Regulation of sinefungin biosynthesis by the wild-type strain and mutants of Streptomyces incarnatus. Appl. Environ. Microbiol., 54:1109-1111 Malina, H. & M.Robert-Gero (1990). Involvement of plasmids in S. incarnatus phenotype. FEMS Microbiol. Lett., 68:195-200 Malina, H. & M.Robert-Gero (1992). Characterization of an 8.7-kilobase thiostrepton resistance-encoding plasmid (pGIF3) of Streptomyces incarnatus. Appl. Environ. Microbiol., 53, (in press )
Malpartida, F., & D.A. Hopwood (1984). Molecular cloning of the whole biosynthetic pathway of a Streptomyces antibiotic and its expression in a heterologous host. Nature, 309:462-464 Maniatis, T.E.F.Fritsch, & J.Sambrook (1982). Molecular cloning. A Laboratory Manual. Cold Spring Harbor Lab.Press, Cold Spring Harbor, New York Murakami, T., C.Nojiri, H.Toyama, E. Mayashi, Y.Yanada & K.Nagaoka (1983). Pock-forming plasmids from antibiotic-producing Streptomyces. J.Antibiot., 36:429434 Murakami,T., H. Anzai, S.Imai, A.Satoh, K. Nagaoka & C.J.Thompson (1986). The bialaphos biosynthetic genes of Streptornyces hygroscopicus: Molecular cloning and characterization of the gene cluster. Mol. Gen. Genet., 205: 42-50 Murray, N.E., W.J.Brammar & K.Murray (1977). Lamboid phages that simplify the recovery of in vitro recombinants. Mol. Gen.Genet., 150:53-61 Parry, R.J., I.YArzu, S. Ju & B.J.Baker (1989). Biosynthesis of sinefungin: on the mode of incorporation of L-ornithine. J.Am. Chem . Soc., 111:8981-8982 Thompson, C.J.J.M.Ward & DA.Hopwood (1982). Cloning of antibiotic resistance and nutritional genes in Streptomyces. J. Bacteriol., 151:668-677 Vedel, M., F.Lawrence, M.Robert-Gero & E. Lederer (1978). The antifungal antibiotic sinefungin as a very active inhibitor of methyltransferases and of the transformation of chick embryo fibroblasts by Roux sarcoma virus. Biochem. Biophys. Res. Comm., 85: 371-376. Copyright 1992 CETA
The following images related to this document are available:Halftone images[ac92006b.gif]Photo images[ac92006b.jpg]Line drawing images[ac92006a.gif] [ac92006c.gif] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}