 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
STRUCTURAL STUDIES OF MAJOR GLYCOLIPIDS FROM CURTOBACTERIUMM FLACCUMFACIENS PATHOVAR BETAE AND ROTHIA DENTOCARIOSA H.MORDARSKA, A.GAMIAN and I.EKIEL * Institute of Immunology and Experimental Therapy, Polish Academy of Sciences, Czerska 12, 53-114 Wroclaw, Poland and * Biotechnology Research Institute, National Research Council, 6100 Royalmount Av., Montreal, Quebec H4P 2R2, Canada
Code Number: AC92007
Sizes of Files:
Text: 20K
Graphics: Line Drawings (gif) 22K
ABSTRACT. Two major glycolipids of Curtobacterium
flaccumfaciens pathovar betae (a, b) and one of
Rothia dentocariosa (c)were isolated, purified and studied
by analytical methods, thin-layer chromatography (TLC),
gas-liquid chromatography (GLC), mass spectrometry (GC-MS) and
nuclear magnetic spectroscopy (NMR). The only components
detected in the samples were mannose, glycerol and fatty
acids. The sugar moiety of glycolipid a was a trisaccharide
and that of glycolipids b and c a disaccharide. The most
characteristic components of the two glycolipids detected in
the cells of C.flaccumfaciens pathovar betae
were cyclopropane fatty acid C17:0 and anteiso
branched chain components C15:0 and C17:0. The only major
fatty acid found in the glycolipid of R.dentocariosa
was anteiso acid C17:0. The three glycolipids seem
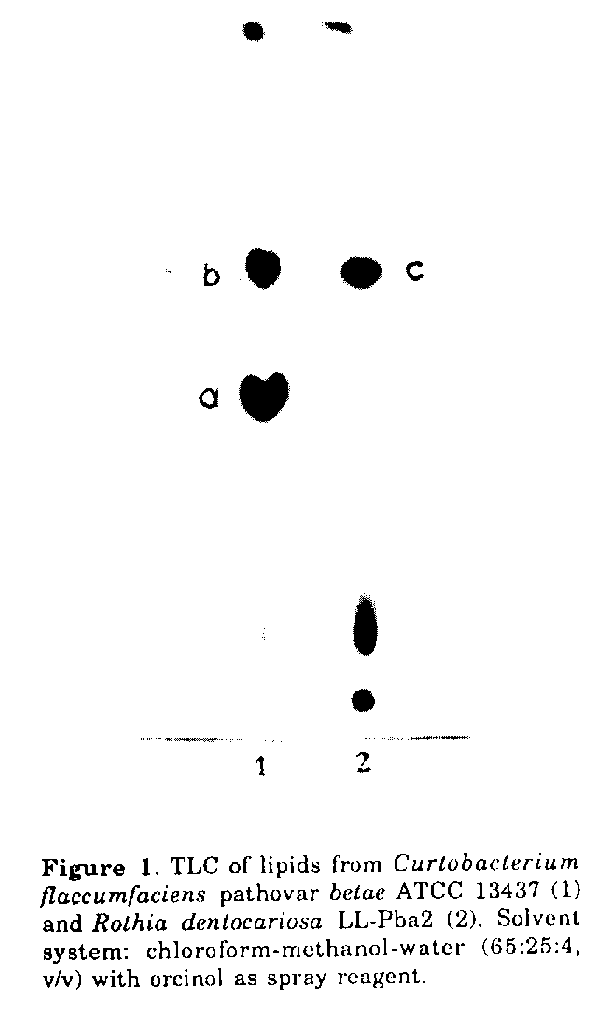
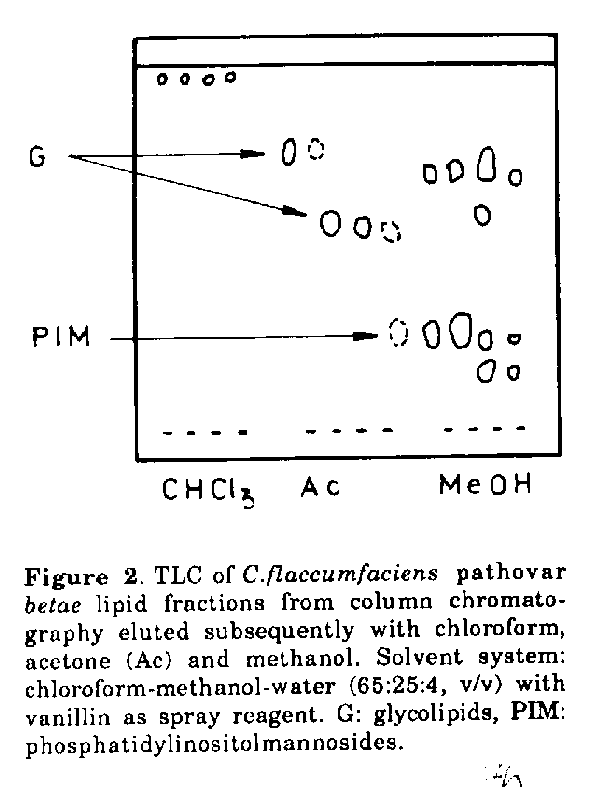
to represent an uncommon structure.Some actinomycetes and allied taxa contain major amounts of glycolipids, which may be useful diagnostic markers in the differentiation of the microorganisms (Mordarska, 1985/86). The present paper deals with some structural studies and comparison of major glycolipids isolated from a plant pathogen Curtobacterium flaccumfaciens pathovar betae, which causes vascular wilt and leaf spot of red beet, and from an opportunistic human pathogen Rothia dentocariosa (Jones and Collins, 1986). The comparable patterns of glycolipids of these bacteria may be considered of taxonomic significance. MATERIALS and METHODS Microorganisms and culture conditions. C.flaccumfaciens pathovar betae (formerly Corynebacterium betae) ATCC 13437 and R.dentocariosa LLPba2 were obtained from the Lechevaliers' Collection, Rutgers University, Waksman Institute of Microbiology, New Jersey, U.S.A. The bacterial strains were grown on yeast extract-dextrose medium in shake cultures as described before (Mordarska, 1985/86). Extraction of lipids. The biomass of the two strains was extracted directly after harvesting using a modification of the Bligh and Dyer procedure (Kates, 1986). Wet cells were twice extracted with chloroform-methanol (2:1, v/v) at 30 C for 12 hrs (water was not added either to the cells or to the mixture of the solvents). The dried lipid extracts were suspended in chloroform-methanol-water (5:5:4.5 v/v) to remove non lipid material. Chromatography. The lipid extracts (ca. 500mg) were fractionated on a column (1.8x45.0cm) of activated Hl-Flosil (60/200 mesh,' Appl.Sci. Lab. Inc., U.S.A.) using reagent grade chloroform (100mlx4), acetone (100mix4) and methanol (100mlx4). The column fractionated samples were monitored for glycolipids by TLC on silica gel H (0.5mm thickness) using the solvent system chloroform-methanol-water (65: 25:4, v/v). Preparative TLC of the acetone eluates containing glycolipids was accomplished by developing silica gel PF 254 plates (20x20 cm) carrying 30 to 40mg of lipids per plate. Glycolipids were detected using a UV lamp and marker components were scraped from plates leaving a margin that was developed with orcinol (Skipski and Barclay, 1969). Bands of orcinol-positive lipids were eluted with reagent grade chloroformmethanol (2:1, v/v) and chloroformmethanol-water (1:1:0.2, v/v). The eluates were brought to dryness under a stream of nitrogen. Preparations were finally purified by rechromatographing samples for up to 3 times (depending on the degree of contamination) as described above. The samples were then checked for purity by two-dimensional chromatography using the solvent system chloroform-methanol-water (65:25: 4, v/v) in one direction and chloroformacetic acid-methanol-water (80:18:12:5, v/v) in the other. Spots detected with vanillin (0.5g dissolved in 100ml of 3% solution of sulphuric acid in ethanol) and orcinol (Skipski and Barclay, 1969) were monitored for phosphorus and ninhydrin (Lechevalier and Lechevalier, 1980). Analytical methods. The total neutral sugar content was measured using the phenol-sulphuric acid method (Dubois et al., 1956) and the phosphorus content after Chen et al. (1956). Solutions were concentrated below 40 C under diminished pressure in a rotary evaporator. Sugar and fatty acids analysis. Samples (0.5mg) of each lipid fraction were hydrolyzed in 0.5ml 1M HCl for 4 hrs at 100 C in sealed tubes. Following evaporation of HCl, samples were reduced and peracetylated according to the procedure of Sewardeker et al. (1965). Analyses were performed on a Packard 5985 B gas chromatographmass spectrometer (GC-MS) using a 3% OV17 capillary column at 180 to 250 C for 6 C/min; an ionization potential of 70eV was applied. Samples (0.4mg) were methanolysed in 1ml of 1M HCl/MeOH for 12 hrs at 80 C for analysis of the fatty acids. Evaporated samples were extracted with chloroform and subjected to analysis using GC-MS under the conditions described above. Methylation analysis. The glycolipids were methylated with methyl iodide in the presence of methyl sulfinyl anion according to the method of Hakomori (1964). The methylated products were recovered by using Sep-Pak C18 car- tridges. Permethylated products were hydrolyzed using 0.5N H2SO4 in 95% CH3COOH (80 C, 18hrs). Methylated sugars were reduced with sodium borodeuteride, acetylated with pyridine- acetic anhydride (1:1, v/v) for 1 hr at 100 C and analyzed by GC-MS using a glass column (2x180mm) with 3% OV17 capillary column on Chromosorb W. The permethylated oligosaccharides were also analyzed directly using the above procedure. De-O-acylation. For the analysis of the sugar moieties the g|ycolipid samples (100p,g) were subjected to alkaline hydrolysis in 5% NH4OH solution for 24 hrs at room temperature. The resultant solution was neutralized with acetic acid, freeze dried and extracted with chloroform-water (1:1, v/v). The aqueous phase was dried in vacuo, dissolved in a minimum volume of water then spotted onto a thin layer silica gel plate. TLC of sugars was carried out on DCFertigplatten Kieselgel 60 F254 (0.25mm) using the solvent system ethyl acetatepyridine-acetic acid-water (5:5:1:3), v/v). The chromatograms were stained with 10% H2SO4 in ethanol. Determination of sugar configuration. A previously described enzymic procedure was used (Gamian et al., 1985). The glycolipid samples (0.4mg) were hydrolyzed (1M HCl, 4 hrs, 100 C) and, once the removal of HCl had been achieved by evaporation, the hydrolyzate was treated using hexokinase (EC 2.7.1.1; Sigma) in the presence of ATP in 0.2M ammonium acetate buffer at pH 8.0. Mannose-6-phosphate was identified by high voltage electrophoresis. NMR spectroscopy. NMR spectra were measured in D2O or benzene d6:methanol-d4 (7:1, v/v) at room temperature using Bruker MSL-300 and AM-360 spectrometers. 13C-NMR signals were assigned using distortionless enhancement by polarization transfer (DEPT) technique (Pegg et al., 1982) and selective proton decoupling. The signals from the 1H-NMR spectra were assigned and the assignment verified by simulation using a Panic program from the Bruker software package. RESULTS Three major glycolipids (Fig. 1) were isolated from the test strains, purified (Fig. 2) and labelled C.flaccumfaciens pathovar betae glycolipids a and b and R. dentocariosa glycolipid c. The glycolipid fractions, which were orcinol and vanillin positive and phosphorus negative, accounted for approximately 0.2% of the wet cell mass.
Figure 2. TLC of C.flaccumfaciens pathovar betae lipid fractions from column chromatography eluted subsequently with chloroform, acerone (Ac) and methanol. Solvent system: chloroform-methanol-water (65:25:4, v/v) with vanillin as spray reagent. G: glycolipids, PIM: phosphatidylinositolmannosides.
The results of the quantitative analyses of the glycolipids are presented in Table 1. The molar ratio of mannose to glycerol was 3:1 in glycolipid a and 2:1 in glycolipids b and c. The presence of mannose was detected by TLC and GLC and confirmed by proton NMR spectra. Analysis of the proton NMR spectra revealed that the coupling constants of mannose were the same (1.7-1.9Hz) in all of the glycolipids studied (Table 2). The configuration of mannose was performed using an enzymic method. Thus, after hydrolysis of the pure samples, mannose was phosphorylated by hexokinase. The presence of mannose-6phosphate identified in high voltage electrophoresis, pointed to the sugar having the D-configuration. The H- and 1C-NMR spectra of glycol1pids b and c were similar suggesting polar head identity. It was evident from lH- and 13C-NMR experiments that the mannose residues had alpha-configuration in glycolipids a, b and c.
--------------------------------------------------------------
GLYCOLIPID
CHARACTERISTIC ---------------------------------------
a b c
----------------------------------------------------------
----
Rf value 0.45 0.63 0.63
Total sugar (%) 62.9 49.1 47.6
Mannose M.R.* 2.56 (3) 2.09 (2) 2.05 (2)
Glycerol MR. 1 1 1
Fatty acids:
ai 15:0 16.4 31.7 0
i 16:0 11.3 4.8 0
n 16:0 10.4 20.1 9.5
ai 17:0 42.3 14.8 45.0
n 18:0 0 0 24.0
/_\ 17:0 9.4 21.5 0
/_\ 19:0 trace trace 0
-------------------------------------------------------------
* Molar ratio (M.R.) of mannose is referred to an arbitrarily
taken single giycerol residue; ai: anteiso, i: iso, n:
normal, A: cyclopropane.Table 1. Chemical analysis of Curiobacterium flaccumfaciens pathovar betae (a and b) and Rothia dentocariosa (c) glycolipids
--------------------------------------------------------------
GLYCOLIPID H-1a* H-1b H-1c H-2a H-2b H-2c
----------------------------------------------------------
----
a 5.06 4.83 4.81 3.97 3.86 4.09
(1.7) (1.7) (1.7)(1.7,3.5)(1.6,3.4)(1.6,3.1)
b 5.09 4.77 3.97 4.08
(1.9) (1.7) (1.7,3.3)(1.9,3.1)
c 5.09 4.77 3.97 4.8
(1.7) (1.7)
-------------------------------------------------------------
* Chemical shifts are in ppm relative to internal
tetramethylsilane. Coupling constants in Hz are given in
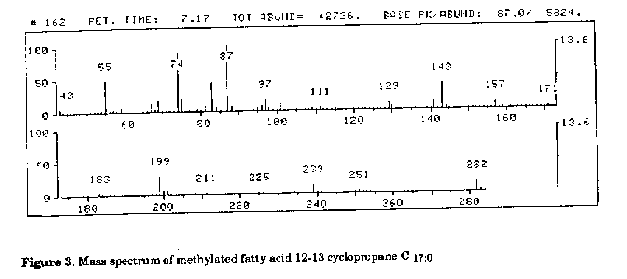
parentheses.Table 2. 1H-NMR results of Curtobacterium flaccumfaciens pathovar betae (a and b) and Rothia dentocariosa (c) glycolipids The methylation analysis indicated that all three glycolipids contained one terminal mannose and 3-substituted mannose. The characteristic components present in both glycolipid fractions of C. flaccumfaciens 'pathovar betae (a, b) were cyclopropane fatty acids. The mass spectrum (Fig. 3) contained characteristic ions: 199, 211,225, 239 and also 55, 69. Molecular ion of m/e = 282 together with gas chromatographic mobility provided evidence that this was a C17:0 cyclopropane fatty acid with the methylene group at the 12 and 13 positions. The other one, C19:0 cyclopropane fatty acid was revealed in trace amounts.
It can be concluded that the major glycolipids of C.flaccumfaciens pathovar betae were different with respect to the amount of sugar present as well as fatty acid composition. The most polar glycolipid a, contained three molecules of mannose and the less polar b, only two. The cyclopropane fatty acids %:0 and the branched anteiso fatty acid were predominant in both the a and b glycolipid fractions. One of the branched-chain fatty acids anteiso-C17:0, was also found in the R.dentocariosa glycolipid c. In the glycolipids studied other normal straight-chain saturated fatty acids were also present. DISCUSSION It can be assumed from the structural studies that g|ycolipids b and c are dimannosyl glycerides and glycolipid a, a trimannosyl glyceride; they have cyclopropane and branched iso and/or anteiso fatty acids, respectively, as predominant components. Fatty acids with branched methyl chains located in ultimate (iso) or penultimate (anteiso) positions are known to make bacteria unique, because they are not common elsewhere (Lechevalier, 1977). They do not seem to occur concurrently with the unsaturated fatty acids as they may serve the same purpose of regulation of membrane fluidity (Jantzen and Bryn, 1985). The results concerning R.dentocariosa glycolipid c are in agreement with the tentative formula reported by Pandhi and Hammond (1975) as dimannosyl digliceride with predominant anteiso C17:0 and a normal C18:0 fatty acids. The glycolipid pattern of C. flaccumfaciens pathovar betae seems to be in agreement with the results of polar lipid analysis (Collins et al., 1980) and nucleic acid hybridization studies on curtobacteria (D6pfer at al., 1982). Some unidentified glycolipids found in "Corynebacterium betae", "Corynebacterium flaccumfaciens", "Corynebacterium poinsettiae" and "Corynebacterium oortii" seem to be very similar to glycolipids a and b. Moreover, the results of the DNA-rRNA cistron similarity, show that the taxa constitute a single homogenous cluster and all of them have presently been reduced to pathovars of one species - Curtobacterium flaccumfaciens (Jones and Collins, 1986). The glycolipid b of C.flaccumfaciens pathovar betae has the same Rf mobility as the glycolipid c from R.dentocariosa. These glycolipids appear to have an identical hydrophilic moiety which consists of one terminal a-D-mannose and 3 substituted a-D-mannose. The total fatty acid composition of the two glycolipids mentioned above and having the same Rf mobility, apart from the predominant anteiso fatty acid C17:0, is different. It seems that distinct specificity and Rf mobility of glycolipids depend mainly on the sugar moiety, which can be easy monitored using TLC with orcinol as spray reagent. The uncommon structure of the glycolipid a (trimannosyl glyceride) as well as fatty acid composition (cyclopropane and branched chain iso and anteiso fatty acids) seem to indicate that the glycolipids would be useful markers in the chemotaxonomy of some Curtobacterium and Rothia taxa. REFERENCES Cheil, P.S., T.Y.Toribara & H.Warner (1956). Microdetermination of phosphorus. Analyt, Chem., 28:1756-1758 Collins, M.D., M.Goodfellow & D.E.Minnikin (1980). Fatty acid, isoprenoid quirtone and polar lipid composition in the classification of Curiobacterium and related taxa. J. gen. Microbiol., 118:29-37 Dopfer, H., E.Stackebrandt & F.Fiedler (1982). Nucleic acid hybridization studies on Microbacterium, Curiobacterium, Agromyces and related taxa. J.gen.Microbiol., 128:1697-1708 Dubois, M., K.A.Gilles, J.K.Hamilton, P.A.Rebers & F.Smith (1956). Colorimetric method for determination of sugars and related substances. Analyt. Chem., 28:350-356 Gamian, A., E.Romanowska, A.Romanowska, C.Lugowski, J.Dabrowski & K.Trauner (1985). Citrobacter lipopolysaccharides: structure elucidation of the O-specific polysaccharide from strain PCM 1487 by mass spectrometry, onedimensional and two-dimensional H-NMR spectroscopy and methylation analysis. Eur.J. Biochem., 146:641-647 Hakomori, S.I. (1964). Rapid permethylation of glycolipids and polysaccharides, catalyzed by methylsulfinylcarbanion in dimethylsulfoxide. J. Biochem. (Tokyo), 55:205-208 Jantzen, E. & K.Bryn (1985). Whole-cell and lipopolysaccharide fatty acids and sugars of Gram-negative bacteria. In: M.Goodfellow & D.E.Minnikin (eds.) Chemical Methods in Bacterial Systematics. Academic Press, London, pp. 145-171 Jones, D., & M.D.Collins (1986). Irregular, nonsporing Gram-positive rods. In: P.H.A. Sneath (ed.) Bergey's Manual of Systematic Bacteriology. Williams & Wilkins, Baltimore, Vol. 2, pp. 1261-1434 Kates, M. (1986). Techniques of lipidology Isolation, analysis and identification of lipids. In: R.H.Burdon & P.H. van Knippenberg (eds.) Laboratory Techniques in Biochemistry and Molecular Biology. Elsevier, Amsterdam, Vol. 3, Part 2, pp. 106-107 Lechevalier, M.P. (1977). Lipids in bacterial taxonomy - A taxonomists view. Crit. Rev. Microbid., 5:109-210 Lechevalier, M.P. & H.A.Lechevalier (1980). The chemotaxonomy of actinomycetes. In: A.Dietz & D.W.Thayer (eds.) Actinomycete Taxonomy. Society for Industrial Microbiology, Arlington, VA, Special publication 6, pp. 227-291 Mordarska, H. (1985/86). Taxonomic value of glycolipids of actinomycetes and allied taxa. The Actinomycetes, 19:11-32 Pandhi, P.N. & B.F.Hammond (1975). A glycolipid from Rothia dentocariosa. Arch. Oral. Biol., 20:399-401 Pegg, DT., D.M.Doddrell & M.R.Bendall (1982). Proton-polarization transfer enhancement of a heteronuclear spin multiplet with preservation of phase coherency and relative component intensities. J. Chem. Phys., 77: 2745-2752 Sawardeker, J.S., J.H.Sloneker & A.R Ieanes: Quantitative determination of monosaccharides as their alditol acetates by gas liquid chromatography. Analyt. Chem., 37: 1602-1604, 1965 Skipski, V.P., & M. Barclay (1969). Thin-layer chromatography of lipids. In: J.M.Lowenstein (ed.) Methods in Enzymology. Academic Press, New York, Vol. 14, pp. 530-598. Copyright 1992 CETA
The following images related to this document are available:Line drawing images[ac92007a.gif] [ac92007b.gif] [ac92007c.gif] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}