 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
A SPORANGIATE ACTINOMYCETE WITH UNUSUAL MORPHOLOGICAL FEATURES: STREPTOSPORANGIUM CLAVIFORME SP. NOV. B. PETROLINI, S. QUARONI, P. SARDI, M. SARACCHI and N. ANDRIOLLO * Istituto di Patologia Vegetale, Universita degli Studi, 20100 Milano and * Istituto Guido Donegani ENICHEM S.p.A., Novara, Italy
Code Number: AC92010
Sizes of Files:
Text: 16K
Graphics: Photos (jpg) 139K / Halftones (gif) 426K
ABSTRACT. A sporangiate actinomycete (strain DI = IPV 2852 = NCB 1160), isolated from decomposed leaf litter of Betula alba using an Andersen sampler connected to a sedimentation chamber, was studied. On the basis of its morphological features and cell wall composition, it was placed in the genus Streptosporangium. This organism is believed to belong to a new s.pecies, Streptosporangium claviforme, because of its characteristic production of club-shaped sporangla.
During an investigation, aimed at detecting so-called rare genera of actinomycetes in natural habitats, a technique widely applied in aerobiological studies and suitable for the isolation of actinomycetes (Lacey and Dutkiewicz, 1976) was employed. Decomposed leaf litter of different plants was used as source material. In this paper a sporangial strain with un- usual morphological features is described. MATERIALS AND METHODS Isolation methods. Betula alba leaves, collected 1-2 weeks after fall, were kept wet for some months to promote decomposition. The leaf litter was then dried at room temperature, reduced to a powder, poured into a sedimentation chamber in which it was agitated for 2 minutes and left to sediment for 30 minutes. The air of the chamber was then aspi- rated for 10 seconds by an Andersen sampler and collected on different media. Strain Di (= IPV-2852, NCB 1160) was isolated from a plate of tap water medium containing 25 g of agar per litre (WA25), after a long incubation period (40 dd at 26 C). The isolate was maintained on oatmeal agar slants and incubated at 26 C frozen as a spore suspension in 10% glycerol at -20 C (Wellington and Williams, 1978) or as slope cultures kept at 4 C. Lyophilization was found to be the most suitable method for longterm preservation. Cultural characteristics. Cultural characteristics were determined on WA25, ISP media 2, 3, 4 (Shirling and Gottlieb, 1966), nutrient agar Difco (NA), Czapek solution agar Difco (CA), potato agar (PA), thin potato-carrot agar (TPC) (Higgins et al., 1967), modified Bennett agar (BA) (Jones, 1949) and Emerson agar (EA) (Waksman, 1961). The results were recorded after 7, 14 and 21 dd of incubation at 26 C using the methods recommended by Shirling and Gottlieb (1966). The extent of growth, colour of both substrate and aerial mycelia, reverse colour of vegetative mycelium, production of soluble pigments and presence of sporing structures were noted. Colour of substrate and aerial mycelia were determined using the colour chart of Prauser (1964). Morphology and fine structure. The morphology of sporing structures was studied by light microscopy and scan- ning electron microscopy (SEM). For SEM observations, small blocks of medium bearing sporing structures were removed from plates and prepared by a previously described technique (Locci and Petrolini Baldan, 1971). Observations were made with a Stereoscan 250 (Cambridge Sci. Instr. Ltd., UK).
--------------------------------------------------------
Substrate myceHum Aerial mycelium
--------------------------------------------- Sporangia
Amount Colour Amount Colour
--------------------------------------------------------
ISP2 (+) Coo5m (hazel) - -
ISP 3 +++ Co6b (pale buff) ++ whitish +
ISP4 + Co6b (pale buff) - -
WA25 ++ Colourless + whitish +
PA + Co5a (pale buff) - -
NA ++ Co58J5b (pale buff)- -
TPC + Co5b (pale buff) - -
BA + Co5b (pale buff) - -
EA ++ Co5a (pale buff) - -
----------------------------------------------------------
Table 1. Cultural characteristics and presence of
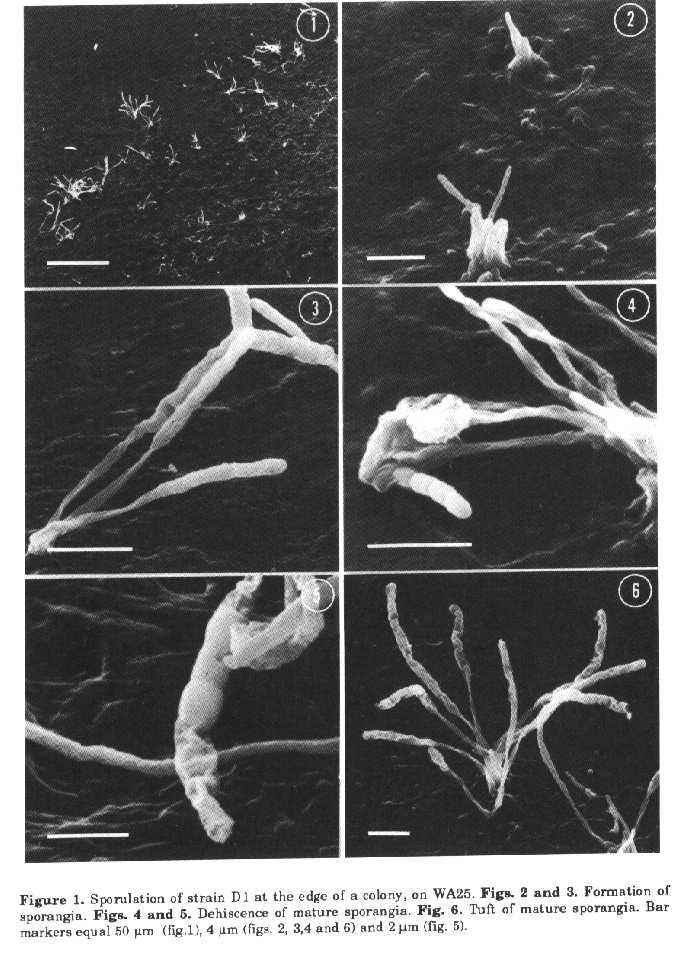
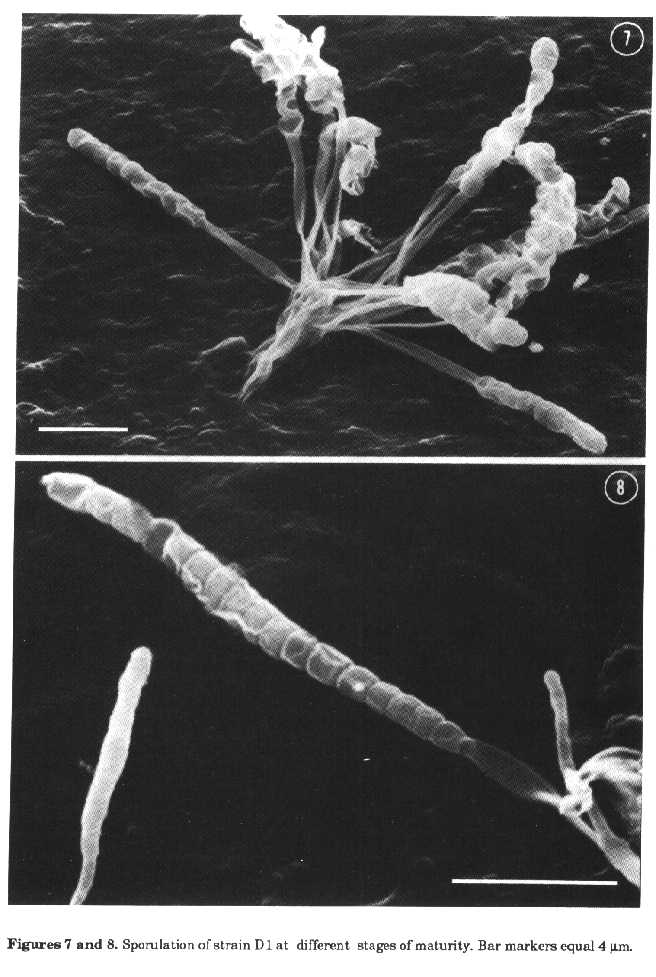
sporing structures of strain DiSporangia suspensions were prepared with TC distilled water (Difco), brain heart infusion (Difco), distilled water with Tween 80 added as a wetting agent and examined by light microscopy to determine sporangia dehiscence and spores motility. Physiological tests. The isolate was incubated on oatmeal agar (ISP medium 3) at a variety of temperatures (4, 10, 26, 37 and 45 C). The following tests were also employed: carbohydrate utilisation on ISP medium 9 and melanin production on ISP media 6 and 7 (Shirling and Gottlieb, 1966), nitrate reduction, H2S production, hydrolysis of gelatine, casein and starch, tyrosine degradation (Williams et al., 1989) and sodium chloride tolerance (3, 4 and 5% NaCl). All the tests were performed at 26 C. Cell chemistry. Mycelium for chemical analyses was obtained from cultures grown on ISP medium 1 (Shirling and Gottlieb, 1966) at room temperature, in flasks on a rotary shaker, harvested by centrifugation, washed and lyophilized. The analyses were carried out as reported by Kutzner (1981). RESULTS The cultural characteristics of isolate DI are shown in Table 1. Substrate mycelium was pale buff and lacked dif- media; the reverse colour of colonies was not distinctive. Aerial growth was whitish and spore differentiation occurred only on ISP medium 3 and WA25, these media being the same used for morphological studies. No growth was observed on CA. Fragmentation of vegetative mycelium did not occur, and spores were formed only on aerial growth. The spore-bearing structures consisted of rudimentary hyphae, gathered into tufts. Soon after isolation, the sporangial tufts were formed on WA25 only at the edge of the colonies and sterile aerial hyphae were not produced (fig. 1); however after a period of cultivation under laboratory conditions sporangia were uniformly distributed over the surface of the agar media. Sterile aerial hyphae were also formed. The tufts consisted of short aerial hyphae emerging from the substrate mycelium and closely bunched together to form a pedestal-like structure bearing sporangia (figs. 2, 4, 6 and 7). The arrangement of sporangia in tufts (generally 3 to 12, sometimes very numerous) is typical for strain D1, nev- ertheless some single sporangia were also found. The sporing structures were clubshaped and when examined at high magnifications, appeared to be covered by a thin envelope. The surface of young sporangia was smooth (fig. 3), then some undulation of the envelope appeared suggesting that spores were present within it (figs. 6 and 7). As the club-shaped structures matured, the envelope became evident because of wrinkling (figs. 7 and 8), then apical dehiscence revealed chains of naked sporangiospores with a smooth surface (fig. 4).
Figures 7 and 8. Sporulation of strain D1 at different stages of maturity. Bar markers equal 4um.
It was also possible to see collapsed sporangial walls, some spores having been released from the sporangium which appeared partially empty (fig. 5). The length of the club-shaped sporangia ranged from 6.5 to 15um (average 10.3um) and sporangiophores appeared to be 5.5+15.5um long (average 7.9um). Each sporangium usually contained 5 to 17 spores in a single straight row. The sporangiospores were subspherical or rod-shaped (0.50+2.0 x 0.65+1.2um) with a smooth surface (figs. 4 and 5). Motile spores were never observed. After 7-14 dd incubation, especially on ISP medium 3, sometimes mature sporangia became bent at their apex or, more rarely, on themselves (figs. 4, 6 and 7). The physiological characteristics of isolate DI are as follows: it grows at temperatures between 10 and 45 C with an optimum between 26 to 37 C; it produces only traces at 10 C and moderate growth at 45 C. It reduces nitrate to nitrite; produces only traces of H2S; does not produce melanoid pigments on ISP media 6 and 7; degrades tyrosine crystals; hydrolyses gelatine, starch and casein; does not grow in the presence of any of the NaCl concentrations utilised. Carbohydrate utilisation is difficult to evaluate because the strain, isolated en a sugar-free medium, grows at the same rate on the basal medium, without any carbon source, and in the presence of glucose, L-rhamnose, meso-inositol and salicin. The growth in the presence of D-lactose, D-mannitol, D-galactose, maltose, raffinose, sucrose, D-xylose, Larabinose, glycerol is greater than that on the control. D-mannose and D-fructose inhibits the development of D1. Cell walls contain meso-diarainopimelic acid but no characteristics sugars. The diagnostic sugar in the wholecell hydrolysates is madurose. Therefore the isolate D1 can be considered as having type III cell walls and the whole-cell sugar pattern B (Lechevalier and Lechevalier, 1970; Lechevalier et al.,1971). DISCUSSION The detection of meso-DAP and madurose is typical of Maduromycetes (Goodfellow, 1989) and of the genera Frankia (Lechevalier and Lechevalier, 1989) and Dermatophilus (Gordon, 1989). The latter are readily distinguished from Maduromycetes on morphological grounds. The generic identification of wall chemotype IIIB sporangiate actinomycetes can be reached using morphological criteria. The isolate D1 produces sporangia with numerous nonmotile spores and, on this account, belongs to the genus Streptosporangium, but has morphological characteristics unusual for the genus. The genus Streptosporangium (Nonomura, 1989) contains actinomycetes producing globose sporangia on aerial mycelium. The nonmotile spores are formed by septation of a coiled unbranched hypha within the sporangium. Club-shaped sporangia have been reported only by Williams and Sharples (1976) for S. corrugatum and, according to these authors, may be regarded as analogous to the spore chains observed in some species of the genus. Nevertheless S. corrugatum forms two types of sporangia, club-shaped and various sized spherical bodies. These later are common, the club-shaped structures are infrequent except in old cultures (21 dd). The species of Streptosporangium have generally a well developed aerial mycelium while aerial hyphae were not usually differentiated in our isolate. For this reason and also because of cultural characteristics strain Di is similar to S. corrugatum. There are, however, some important differences. Isolate DI contains a larger number of spores within the sporangium, does not produce globose sporangia and the surface of its spores is smooth. On the basis of the above mentioned characteristics it is believed that the isolate D1 belongs to a new species, for which the name Streptosporangium claviforme (club-shaped) is proposed. Description of Streptosporangium claviforme sp. nov. Stable branched mycelium is formed with poor aerial hyphae bearing clubshaped sporangia (6.5:15 um, average 10.3 um) containing 5+17 spores in a single straight row. Sporangiophores are long (5.5+15.5 um, average 7.9 um). Sporangia are formed only on aerial mycelium, generally in tufts, and are abundant on WA25 and ISP medium 3. The sporangiospores are subspherical or rod-shaped (0.50+0.65 x 2.0+1.2 um) with a smooth surface, non motile. The substrate mycelium is pale buff, the reverse side of the colony is beige and soluble pigment is not produced. The aerial mycelium, if present, is whitish. Cell walls contain meso-DAP and whole-cell hydrolysates contain madurose. Since the species description is based on a single isolate, strain Di (= IPV 2852, NCB 1160) is the type strain of S. claviforme. The type strain is deposited at the National Culture Bank, Gorizia, as NCB 1160. REFERENCES Goodfellow, M. (1989). Maduromycetes. In: S.T.Williams (ed.) Bergey's Manual of Systematic Bacteriology. The Williams & Wilkins Co., Baltimore, Vol. 4, pp. 2509-2551 Gordon, M.A (1989). Genus Dermatophilus Van Saceghem 1915, 357, emend. mut. char. Gordon 1964, 521AL. In: S.T.Williams (ed.) Bergey's Manual of Systematic Bacteriology. The Williams & Wilkins Co., Baltimore, Vol. 4, pp. 2409-2410 Higgins, M.L., M.P.Lechevalier & H.A. Lechevalier (1967). Flagellated actinomycetes. J. Bacteriol., 93:1446-1451 Jones, K.L. (1949). Fresh isolates of actinomycetes in which the presence of sporogenous aerial mycelia is a fluctuating characteristic. J. Bacteriol., 57:141-145 Kutzner, H.J. (1981). The family Streptomycetaceae. In: M.P.Starr et al. (eds.) The Prokaryotes. Springer Verlag, Berlin, Vol. 2, pp. 2028-2090 Lacey, J. & J-Dutkiewiez (1976). Isolation of actinomycetes and fungi using a sedimentation chamber. JAvpl. Bacteriol., 41:315-319 Lechevalier, M.P. & H.A. Lechevalier (1970). Chemical composition as a criterion in the classification of aerobic actinomycetes. Int. J. Syst. Bacteriol., 20:435443 Lechevalier, M.P. & H.A. Lechevalier (1989). Genus Frankia Brunchorst 1886, 174AL. In: S.T. Williams (ed.) Bergeys Manual of Systematic Bacteriology. The Williams & Wilkins Co., Baltimore, Vol. 4, pp. 2410-2417 Lechevalier, H.A., M.P Lechevalier & N.N. Gerber (1971). Chemical composition as a criterion in the classification of actinomycetes. Adv. Appl. Microbiol., 14:47-72 Locci, R. & B.Petrolini Baldan (1971). On the spore formation process in Actinomycetes. V. Scanning electron microscopy of some genera of Actinoplanaceae. Riv.Pat. Veg., S IV, 7 (Suppl.): 81-96 Nonomura, H. (1989). Genus Streptosporangium Couch 1955, 148AL. In: S.T. Williams (ed.) Bergey's Manual of Systematic Bacteriology. The Williams & Wilkins Co., Baltimore, Vol. 4, pp. 2545-2551 Prauser, H. (1964). Aptness and application of colour codes for exact description of colours of streptomycetes. Z-Allg. Mikrobiol., 4:95-98 Shirling, E.B. & D.Gottlieb (1966). Methods for characterization of Streptomyces species. Int. J. Syst. Bacteriol., 16, 313-340 Waksman, SA. (1962). The Actinomycetes. Classification, Identification and Descriptions of Genera and Species, Vol. 2, The Williams and Wilkins Co., Baltimore Wellington, E.M.H. & S.T.Williams (1978). Preservation of actinomycete inoculum in frozen glycerol. Microbios Lett., 6:151-157 Williams, S.T. & G.P. Sharples (1976). Streptosporangium corrugatum sp. nov., an actinomycete with some unusual morphological features. Int. J. Syst. Bacteriol., 26:45-52 Williams, S.T., M.Goodfellow & GJklderson (1989). Genus Streptomyces Waksman and Henrici 1943, 339AL.In: S.T.Williams (ed.) Bergey's Manual of Systematic Bacteriology. The Williams & Wilkins Co., Baltimore, Vol. 4, pp. 2452-2492. Copyright 1992 CETA
The following images related to this document are available:Halftone images[ac92010a.gif] [ac92010b.gif]Photo images[ac92010a.jpg] [ac92010b.jpg] |
| |||||||||
{kind=link}
{kind=link}