 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
U. GRAFE, W. IHN and I. ERITT Hans-Knoll-Institute of Natural Product Research, Beutenbergstrasse 11, PO Box 100 813, D-07708 Jena, Germany
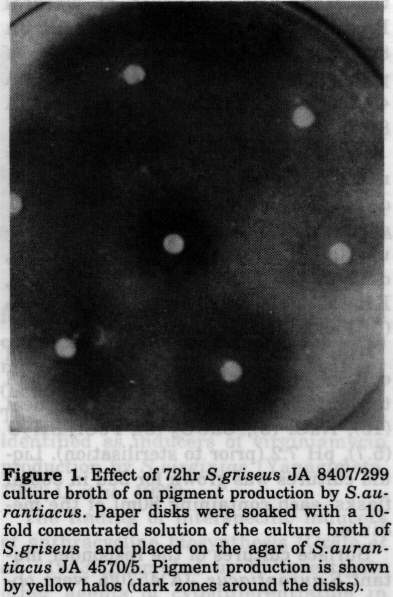
Several representatives of the Actinomycetales have been shown to produce autoregulatory factors which govern the formation of aerial mycelium and secondary metabolites at a very low concentration (Grafe, 1989; Beppu, 1992). An example is provided by the butyrolactone-type A-factor which acts as an inducer of streptomycin and anthracycline biosyntheses in non-sporulating strains of Streptomyces griseus (Kleiner et al., 1976; Eritt et al., 1984). For instance, the addition of a very low amount of this metabolite to growing cultures of the pertinent idiotrophic mutant strain re-established both morphological differentiation (e.g., the formation of aerial mycelium and spores) and the production of antibiotics (Grafe et al., 1984; Horinouchi et al., 1984). Obviously, the A-factor serves as a kind of a microbial hormone representing a key intermediate within a signal chain that triggers secondary metabolism and morphological development (Hara and Beppu, 1982; Horinouchi and Beppu, 1990). Moreover, in our previous work we showed that even dihydro-derivatives of A-factor are produced by a series of different Streptomyces species and induce cytodifferentiation of blocked S. griseus mutants in an interspecific manner (Grafe et al., 1981; Grafe et a1., 1983). The dihydro-derivatives of A-factor were identified as inducers of virginiamycin production by S. virginiae (Yanagimoto, 1983; Li et al., 1992). The finding of interspecific inducers of anthracycline biosynthesis (Grafe et al. , 1983) prompted us to investigate other agents of cytodifferentiation in surface cultures of other streptomycetes . While screening for substances from actinomycete cultures inducing pigment formation in different genera and species, cell-free culture samples of S. griseus mutant 299 were shown to stimulate the formation of a yellow pigment by surface cultures of the mutant strain S. aurantiacus JA 4570/5. Here we report the isolation and the structure elucidation of this antibiotic. A second aim of the present work was to identify the chemical nature of the inducing factor from S. griseus JA 8407/299 exerting an interspecific effect on secondary metabolism of S. aurantiacus JA 4570/5. MATERIALS and METHODS Strains and growth conditions. S. griseus JA 8407, S. aurantiacus JA 4570 and the other strains mentioned were obtained from the culture collection of the Hans-Knoll-Institute of Natural Product Research, Jena. S. griseus JA 8407/299, producing the stimulator of pigment formation by S. aurantiacus JA 4570/5, was a mutant of S. griseus JA 8407. To isolate the inducer of pigment formation affecting surface cultures of S. aurantiacus JA 4570/5, the mutant S. griseus 299 was grown on A1 53 medium (g/l distilled water): sucrose (3), dextrin (15), urea (0.1), NaCl (0.5), KH2PO4 (0.5), yeast extract (1), peptone (5), FeSO4.6H20 (1), agar (15), pH 7.2. The medium for inoculum and liquid culture preparation was as follows (g/l tap water): glucose (30), soya bean meal (10), NaCl (3), CaC03(3), FeCl3.6H20 (5.7), pH 7.2 (prior to sterilisation). Liquid cultures was grown for 3dd in 500ml flasks, each containing 80ml of the medium . Surface cultures of the responder mutant S. aurantiacus JA 4570/5 were obtained by inoculating the strain from agar cultures onto oatmeal-agar medium consisting of (g/l tap water): oatmeal (20), Difco agar (15), pH 7.2 for 10dd. To demonstrate the inducing effect of the S. griseus metabolite(s) on pigment formation, agar wells were cut into freshly inoculated cultures of S. aurantiacus 4570/5 and filled with 100ul of a 72 hrs culture liquid of S. griseus 8407/299. Alternatively, dried paper disks soaked with 100ul of the culture broth of S. griseus were applied to the agar surface. Formation of a yellowish halo around the samples after 2-3dd indicated stimulation of pigment production. Chemicals. All chemicals used were analytical grade. NADH, NAD, NADPH, NADP, ATP, ADP, AMP, GTP, GDP, GMP, CTP, CDP, UTP, TTP, TDP, TMP, cytosine, thymidine, pyridoxal-5'-phosphate, nicotinamide and other reagents mentioned in Table 1 were purchased from Boehringer (Mannheim) and Merck (Darmstadt). Preparative isolation of the yellow pigment. Fifty agar plates (25cm diameter) containing a 0.5cm layer of medium were inoculated with S. aurantiacus JA 4570/5. Agar wells (12 per plate) were filled with 100ul of a cell-free 72hr culture broth of S. griseus 299. After 72hrs at 28 C the whole agar culture was extracted twice with 5l of ethyl acetate. The yellow extract was dried, concentrated and subjected to column chromatography on Sephadex LH-20 (in methanol) and silica gel 60 (Merck, 0.063-0.1um, CHCl3/MeOH, 9:1 v/v). The yellow fractions, displaying an antibacterial effect against Bacillus subtilis ATCC 6633, were pooled and evaporated. Final purification was achieved by HPLC (silica gel RP18], acetonitril/water, 6:4, 2ml H3PO4/500ml; 0.6ml/min detection at 242 nm ) . Yield was 6mg (yellow powder). FT IR Spectrum (in KBr, cm-1) 820, 890, 910, 1085, 1120, 1160, 1185, 1220, 1260, 1290, 1315, 1335, 1350, 1380, 1410, 1450, 1510, 1600, 1622, 1645, 1710, 2920, 2975, 3400. UV spectrum (l[max]): 253nm, 265nm, 420nm. HR-MS (mass spectrometer AMD 402, Harpstedt, Germany): M+ m/z 580.1705 (calcd. 580.1721 for C[34]H[28]0[9]) M+-H20 m/z 562.1616 (calcd. 562.1616 for C[34]H[26]O[9]) M+-2H20 m/z 544.1523 (calcd. 544.1535 for C[34]H[24]0[7]). 150 MHz ^13C-NMR (chemical shift in ppm, CDC[13]): 22.9; 28.1; 29.4; 33.8; 47.1; 60.6; 60.8; 72.1; 108.6; 109.2; 112.4; 112.6; 113.0; 116.4; 117.4; 124.2; 125.0; 125.8; 133.1; 133.5; 133.8; 134.3; 137.0; 138.6; 158.0; 158.5; 158.7; 164.8; 166.4; 191.0; 199.4; 203.1; 288.3 . Isolation of the inducer of setomimycin production. The active principle from 10 litres of a 72hr culture broth of S. griseus JA 8407/299 was adsorbed to Amberlite XAD 1180 (Serva, Heidelberg). It was eluted by washing the resin with methanol/water (6:4), methanol was removed in vacuo, and the aqueous residue applied to the top of a column (3cm x 15cm) filled with DEAESephadex A-20 in water. The active substance was not adsorbed while proteinaceous impurities were retained. The eluate was lyophilised, dissolved in 15 ml of water and subjected to column chromatography on cellulose (Merck, Avicel). The first eluent was methanol, followed by a gradient of MeOH/Water (9:1 to 8:8). The bioactivity of all fractions was tested against S. aurantiacus JA 4570/5, and the positive ones were lyophilised, again. Small portions were dissolved in water and separated by preparative thin-layer chromatography (TLC) using Merck plates (20 x 20cm) precoated with silica gel RP[18] (MeOH/ water, 1:1 ). Two of the strongly fluorescent zones (W 254 nm) on the chromatograms, at Rf 0.9 (inducer A) and 0.7 (inducer B) were scraped off, eluted with the same solvents and Iyophilised. Yield of crude inducer A was 8mg. Further purification was achieved by HPLC (acetonitril/water; 1:1; 0.5ml/ min, retention time 5.3min, Spherisorb RP18, column 4.3 x 250mm), with a yield of 1.5mg. 200 MHz lH NMR (in DMSO, chemical shift in ppm): 1.0; 1.05; 1.1; 3.5; 4.1; 7.1; 7.4. RESULTS and DISCUSSION The stimulating effect of the culture liquid of S. griseus mutant 299 on the formation of a yellow pigment by a 3dd surface culture of the S. aurantiacus mutant JA 4570/5 is shown in Fig. 1. Figure.1 Effect of 72hr S. griseus JA 8407/299 culture broth of on pigment production by S.aurantiacus. Paper disks were soaked with a 10fold concentrated solution of the culture broth of S. griseus and placed on the agar of S. aurantiacus JA 4570/5. Pigment production is shown by yellow halos (dark zones around the disks).
The S. griseus strain does not produce the yellow pigment
and surface cultures of the parental strain S. aurantiacus
JA 4570) turn yellow without the inducer, but only after 8-lOdd
growth at 25 C. Obviously the effective substances secreted by
S. griseus do not affect S. aurantiacus JA 4570/5 as
an inducing factor, but appear to act as a kind of a stimulator
determining an earlier onset of secondary metabolite production.
Approximately 15% of a series of Streptomyces strains from
our culture collection and belonging to different taxonomic groups,
namely, strains JA 8354, JA 8356, JA 8362, JA 8364 and JA 8799,
display the same inducing effect on pigment production.
The yellow pigment, isolated from surface cultures of S.
aurantiacus JA 4570/5, analysed by mass spectrometry and
high-field NMR, proved to be identical with the antitumor
antibiotic setomimycin (I, Figs. 2 and 3) (Omura et al.,
1978; Kakinuma et al., 1981). Moreover, the molar mass
of the pigment was estimated by FAB-mass spectrometry as 580 Da
(M+H^+, m/z 581; C[34]H[29]0[9]).
Figure 3. 400 MHz-1H, 1H-COSY spectrum of setomimycin isolated
from agar cultures of S. aurantiacus (in CDCl[3];
chemical shift in ppm).
Figure 4. Fluorescence spectra of the stimulator of
setomimycin production isolated from broth cultures of S
griseus (circles), and of NADH (crosses) (in water).
Shaded symbols represent samples after the addition of sodium
pyruvate and lactate dehydrogenase.
Setomimycin (I) is a typical acetogenine metabolite and its
biosynthesis by S. pseudovenezuelae was shown to occur
via the polyketide route, employing only acetate-derived
building blocks (Kakinuma et al. , 1981).
In order to investigate the structure of the stimulating
factor produced by S. griseus strain 299, active substances
from liquid cultures were enriched by XAD 1180 adsorber resin,
removal of proteinaceous impurities by ion exchange chromatography
on DEAE-Sephadex A-207 column chromatography on cellulose and
reverse phase silica gel RP[18] chromatography. The inducer(s) were
rather unstable, and complete purification could not be achieved.
The TLC spots possessing stimulatory activity on pigment production
(at least 0.4 mg per paper disk were required) display strong
fluorescence (maximum at 447nm) under UV light (excitation at
254nm). This finding suggest that pyrimidine coenzymes could
constitute at least one of the inducers of setomimycin production
present in broth cultures of S. griseus 299. The
fluorescence spectrum of zone A was proved to be identical with
that of reduced nicotineamide-adenine dinucleotide (NADH II; Fig.4
.
Figure 5. Structures of several stimulators of setomimycin
production by S. aurantiacus. II: NADH (R= H), III:
NADPH (R=PO3H2), IV: AMP (R=PO3H2), V: ADP (R=P03HPO3H2), VI:
ATP (R= PO3HPO3HPO3H2), VII: GMP (R=PO3H), VIII: GDP
(R=PO3HPO3H2), IX: GTP ( R=PO3HPO3HOP3H2), X: pyridoxal
5'-phosphate (P: PO3H2).
In addition fluorescence disappeared when pyruvate and lactate
dehydrogenase were added to the aqueous solution of zone A (Fig. 4
. Its identity with coenzyme NADH (II) was further supported by
the characteristic proton signals found in the 200 MHz ^1H NMR
spectrum of the partially purified sample. Zone B from the above
chromatograms displayed an Rf similar to NADPH (II) but the scarce
amounts isolated, did not allow further identification.
A final proof in favour of the suggestion of a stimulatory
role played by pyridine coenzymes in setomimycin production by
S. aurantiacus JA 4570 was provided by the addition of
commercial NADH (II) and NADPH (III) to agar plate cultures of the
indicator strain, S. aurantiacus JA 4570/5. Amounts higher
than 50 ug of both NADH and NADPH brought about the same effect on
setomimycin production as the culture broth of S. griseus,
with the size of the halos depending on the concentration.
However the oxidised coenzymes NAD^+ and NADP^+ displayed the
same effect suggesting that NADH and NADPH do not supply reducing
equivalents for setomimycin biosynthesis.
This result led to a systematic investigation of the role of
molar constituents of NAD(P)(H) and of other coenzymes, so as to
characterise the stimulatory molecules. Both nicotinamide and
D-ribose, as major constituents of NAD(P)(H), were inactive in
comparable concentrations but several adenosine phosphates (IV, V,
VI) showed a positive effect. Obviously the adenine moiety of
NAD(P)(H) was not the only inducer, since guanosine (VII, VIII, IX)
and inosine phosphates were also active within the same range of
concentration (Fig. 5).
However, when a series of vitamins and trace elements were
tested, pyridoxal 5' phosphate (X) was shown to exert the same
effect as the purine coenzymes .
It is remarkable that of all the pyrimidine coenzymes tested,
both phosphorylated and non-phosphorylated, not one showed any
stimulatory activity (Table I).
-------------------------------------------------------------------
INDUCER EFFECT
(100 ug)
-------------------------------------------------------------------
---
In order to explain the stimulation of setomimycin biosynthesis by
purines and pyridoxal 5'-phosphates two questions could be raised:
what is the role of purines in setomimycin production and why does
setomimycin formation by S. aurantiacus JA 4570/5 depend on
these factors? At first sight, no direct role of purines in
setomimycin biosynthesis could be recognised, e.g., as
carbon precursors of the carbocyclic skeleton (Omura et al.,
1978; Kakinuma et al., 1981). Possibly, purines and
pyridine coenzymes such as NAD+, NADP+, NADH, NADPH could be
required for special steps of precursor activation and redox
transformations of the biosynthetic intermediates (McCormick and
Morton, 1982), but no details of the early steps of setomimycin
biosynthesis have so far been published (Omura et al., 1978;
Kakinuma et al., 1981). The responder strain S.
aurantiacus JA 4570/5 thus appears as a derivative of the
parental strain S. aurantiacus JA 4570 (Grafe et al.,
1993) producing only a reduced amount of purines and NAD(P)(H)
during the early growth phase. This amount seems to be insufficient
to support both the mycelial development and secondary metabolism.
Even in the parental strain JA 4570 the production of setomimycin
starts after 810dd when growth of surface cultures ceases. Hence it
could be hypothesised that excessive formation of purines plays a
role as a regulatory signal and inducer of secondary metabolism and
cytodifferentiation, at least in strain JA 4570/5 of S.
aurantiacus .
On the other hand, the stimulating effect of pyridoxal
5'-phosphate on setomimycin production by S. aurantiacus JA
4570/5 could be interpreted in terms of metabolic imbalance of
purine biosynthesis in the mutant strain. In contrast to the
formation of pyrimidines, purine production depends on a series of
transaminating enzymes such as, e.g.,
phosphoribosyl-pyrophosphate amidotransferase,
phosphoribosylphosphate-glycinamide-synthetase, and
N-formylglycineamidine-ribonucleotide amidotransferase which all
require this coenzyme (Stryer, 1992). However, during growth of the
young mycelia on agar surfaces, the endogenous level of pyridoxal
5' phosphate could be growth-limiting in S. aurantiacus JA
4570/5, maybe because of specific, genetically determined,
metabolic conditions of the strain .
Finally an interesting point for discussion and future
investigation is that extracellular formation of purines and
pyrimidine coenzymes by Streptomyces can be detected by
means of S. aurantiacus JA 4570/5 as a specific indicator
strain responding to the coenzymes by improved pigment production.
It also appears worthwhile clarifying the general role of
extracellular purines in the physiology and mycelial development of
actinomycetes .
ACKNOWLEDGEMENTS. We gratefully acknowledge the kind assistance of
Prof. Dr. A. Zeeck (University Gottingen, Institute of Organic
Chemistry) in recording the NMR spectra of the yellow pigment.
REFERENCES
Beppu, T. (1992). Secondary metabolism as chemical signals for
cellular differentiation. Gene 115: 159-165
Eritt, I., U. Grafe & W. F. Fleck (1984). Inducers of both
cytodifferentiation and anthracycline biosynthesis of
Streptomyces griseus and their occurrence in actinomycetes
and other microorganisms. Allg. Mikrobiol., 24: 3-11
Grafe, U. ( 1989). Autoregulatory secondary metabolites. In:
S.Shapiro (ed.) Regulation of Secondary Metabolism in
Actinomycetes. CRC Press, Boca Raton, Florida, pp. 75- 126
Grafe, U., I. Eritt & W. F. Fleck (1984). On the role of
A-factor in cytodifferentiation of anthracycline-producing strains
of Streptomyces griseus. Biol. Actinomycetes Rel. Org., 18:
220-136
Grafe, U., G. Reinhardt, I. Elitt, W. Schade, W. F. Fleck & L.
Radics (1983). Interspecific inducers of cytodifferentiation
and anthracycline biosynthesis in Streptomyces bikiniensis
and S. cyaneofuscatus. Biotechnol. Lett., 5: 591-599
Grafe, U. W. Schade, I. Eritt, W. F.Fleck & L. Radics
(i981). A new inducer of anthracycline biosynthesis from S.
uiridochromogenes. J.Antibiot. , 35: 1722-1724
Grafe, U., C. Stengel, R. Schlegel, K. Dornberger, W. Ihn, M.
Ritzau & W. Gunther (1993) Isolation and structure of
pamamycin621. Nat. Prod.Lett. (in press)
Hara, O. & T.Beppu (1982). Mutants blocked in streptomycin
production in Streptomyces griseus - the role of A-factor.
J. Antibiot. , 35: 349-358
Horinouchi, S., Y. Kumada & T. Beppu (1984). Unstable
genetic determinant of A factor biosynthesis in
streptomycin-producing organisms: cloning and characterization.
J. Bact., 158: 481-492
Horinouchi, S. & T. Beppu (1990). Autoregulatory factors of
secondary metabolism and morphogenesis in actinomycetes. CRC
Crit. Reu.Biotechnol., 10: 191-204
Kakiunuma, K., N. Iwamura, N. Ikekawa, H. Tanaka, S.Minami &
N.Ouma (1981). Structure and biosynthesis of setomimycin, a novel
9,9'-bianthryl antibiotic. J. Amer. Chem. Soc., 102:
7493-7498
Kleiner, E. M., S. A. Pliner, V. S. Soifer, V. V.
Onopri,enko, T. A. Balashova, B. V. Rozow & A. S. Khohklov
(1974). Structure of the A-factor, a bioregulator from
Streptomyces griseus. Bioorg. Khimia (Moscow), 2: 825-833
Li, W., T. Nimura, S. Sakuda, T. Nishida & Y. Yamada (1992). New
inducing factors for virginiamycin production from Streptomyces
antibioticus. J. Ferm. Bioeng., 74: 214-217
McCormick, J. R. D. & G. O. Morton (1982). Identity
of the cosynthetic factor I of Streptomyces aureofaciens and
fragment FO from coenzyme F420 of Methanobacterium species.,
J. Amer. Chem.Soc., 104: 4014-4023
Omura, S., H. Tanaka, Y. Iwai, K. Nishigaki, J.Awaya,
Y.Takakashi & R.Masuma (1978). A new antibiotic, setomimycin,
produced by a strain of Streptomyces. J.Antibiot., 31: 1091-
1098
Stryer, L. (1992). Biochemistry. Spektrum
Akademischer Verlag, Heidelberg
Yanagimoto, M. (1983). Physiological studies on
staphylomycin production by Streptomyces virginiae. Novel
actions of inducer of staphylomycin production by Streptomyces
virginiae. J.Ferm. Technol., 61: 433-455.
Copyright 1994 C. E. T. A.
|
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}