 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
THE MAJOR GLYCOLIPID FROM SACCHAROPOLYSPORA HIRSUTA: TAXONOMIC SIGNIFICANCE AND PRELIMINARY CHEMICAL STUDIES H. MORDARSKA and A. GAMIAN Institute of Immunology and Experimental Therapy, Polish Academy of Sciences, Czerska 12, 53-114 Wroclaw, Poland
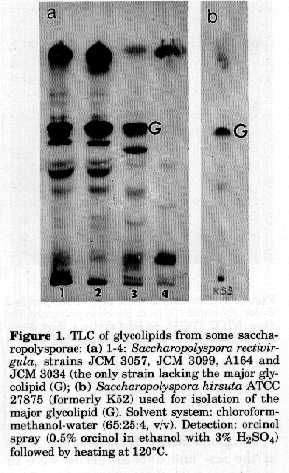
ABSTRACT. A major glycolipid characterised by the same TLC mobility was found in most Saccharopolyspora but not in other genera of the family Pseudonocardiaceae. This glycolipid was isolated from the type strain of Saccharopolyspora hirsuta, purified and examined by analytical methods, gas-liquid chromatography and mass spectrometry. The components detected in the glycolipid were mannose, glycerol and the branched fatty acids anteiso C17:0, iso C17:0, iso C16:0. The sugar moiety of the glycolipid was a disaccharide where one of the fatty acids was shown to be attached to the internal mannose. This uncommon dimannosyl monoacylglyceride might be a valuable marker in the differentiation of Saccharopolyspora from allied taxa. During earlier investigations on glycolipids of actinomycetes and related organisms (Mordarska, 1985/1986) a major glycolipid with a similar TLC behaviour was found in Saccharopolyspora hirsuta and Saccharopolyspora rectivirgula (formerly Faenia rectivirgula). Goodfellow et al., (1989) analysed the polar lipids from Saccharopolyspora gregorii and Saccharopolyspora hordei by TLC and found some uncharacterised glycolipids one of which seems to be chromatographically similar to that mentioned above. These observations, as well as reclassification of Faenia rectivirgula into the genus Saccharopolyspora as Saccharopolyspora rectivirgula (Korn-Wendisch et al., 1989), prompted us to test some additional actinomycetes for the major glycolipid, especially among the members of the new taxa recently included in the family Pseudonocardiaceae (Embley, 1992). It seemed also interesting to undertake some chemical studies on the glycolipid from the type strain of Saccharopolyspora hirsuta, type species of the genus. MATERIALS and METHODS Organisms and growth conditions. All strains investigated and other previously analysed, are listed in Table 1. The strains were grown on yeast extract-dextrose medium in submerged cultures as previously described (Mordarska, 1985/ 1986). Extraction of lipids. Crude lipids were obtained directly from wet cell mass by extraction with chloroform- methanol (2:1, v/v). The dried extract from the type strain Saccharopolyspora hirsuta (chosen for the isolation of the major glycolipid) was suspended in chloroform-methanol- water (5:5:4.5, v/v) to remove water soluble contaminants (Kates, 1986). Thin-layer chromatography (TLC) of the crude lipids. Samples from different taxa of the family Pseudonocardiaceae were analysed by TLC as previously detailed (Mordarska, 1985/1986). Isolation and purification of the major glycolipid from Saccharopolyspora hirsuta. The lipid extract (500mg, free of non lipid material) was fractionated on a column (1.8 x 45cm) of activated silica gel 60 (Merck, 70-230 mesh) using reagent grade chloroform (100ml x 4), acetone (100ml x 5) and methanol (100ml x 4). The eluates were monitored by TLC and all fractions containing the major glycolipid were combined for purification by preparative TLC as previously described (Mordarska et al., 1992). Analytical methods. Total neutral sugar, fatty acid and sugar determinations and methylation analyses were carried out according to Gamian et al. (1993). De-O-acylation. The procedure was performed on a sample (1mg) subjected to alkaline hydrolysis in NH4OH (5% aqueous solution) at 20 C for 3 days. The resultant solution was neutralised with acetic acid, evaporated with a stream of N2 and extracted with chloroform-water (1:1, v/v). For further analyses the oligosaccharide was recovered from the water phase by freeze-drying. Gas liquid chromatography/mass spectrometry. The analysis was carried out with a Hewlett-Packard 5971 A system using an HP-1 (0.2mm x 12m) glass capillary column and temperature program (150-270 C) of 8 /min. Determination of sugar configuration and NMR spectroscopy. The glycolipid sample (0.4mg) was hydrolysed (1M HCl, 4 hrs, 100 C) and released sugar units were treated by hexokinase in 0.2 M ammonium acetate buffer pH 8.0, in the presence of ATP. Mannose-phosphate was identified by high voltage electrophoresis (Gamian et al., 1991). NMR spectra were measured at 100 MHz using a Bruker AM 300 spectrometer.
--------------------------------------------------------------
TAXON STRAINS GLa
AMYCOLATA^b ATCC 19727, IMRU 1105, IMRU 352
3/0
A.autotrophica
AMYCOLATOPSlS^b ATCC 13685, K98
A.mediterranei K99, N857, N858, N87
A.orientalis 6/0
KIBDELOSPORANGIUM
NDc
PSEUDONOCARDIA K24, K88, K125, N752, N756, N761,
P. thermophila LL-A18 LL-SR1b
Pseudonocardia sp. 8/0
SACCHAROMONOSPORA JCM 3098
S.caesia JCM 3315
S.internatus JCM 3036, JCM 3120, Kb (61, 71,
S.viridis 72, 74) 8/0
SACCHAROPOLYSPORA K600
S.erythraea ATCC 27875b*, K16, K51, K55, K57
S.hirsuta JCM 3034d, JCM 3057, JCM 3099, LLb
S.rectivirgula (A91, A94, V4066)
IMRU 1258b, Ab (163, 164), N745
Saccharopolyspora sp.
LL-100-46b 17/16
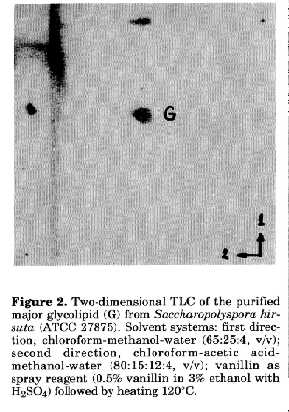
Table 1. Occurrence of the major glycolipid of Saccharopolyspora in taxa of the Pseudonocardiaceae [a Strains studied/strains having the major glycolipid. b Strains previously studied by Mordarska (1985/ 1986); b* Strain previously labelled K52 (Embley et al., 1988a) and used for isolation and chemical studies of the major glycolipid. c Not determined. d The only strain of the taxon lacking the major glycolipid. IMRU: Culture Collection, Waksman Institute of Microbiology, Rutgers University, USA; JCM: Japan Collection of Microorganisms, Tokyo, Japan; A, K, N: strains received from M. Goodfellow, The University of Newcastle upon Tyne, UK; LL: received from H.Lechevalier, Waksman Institute of Microbiology, Rutgers University, USA]. -------------------------------------------------------------- RESULTS and DISCUSSION Occurrence of the major glycolipid from Saccharopolyspora in other taxa of the family Pseudonocardiaceae. Over 40 strains of actinomycetes representing nearly all genera (including Saccharopolyspora) of the family Pseudonocardiaceae were compared for the occurrence of the characteristic major glycolipid detected in Saccharopolyspora hirsuta. Some of them had been previously investigated (Mordarska, 1985/1986). Others, more recently included in the family Pseudonocardiaceae, were analysed during this study (Table 1). It is important to stress that the major glycolipid, found earlier in the strains of Saccharopolyspora hirsuta (Mordarska, 1985/1986), occurred in all tested Saccharopolyspora strains, with the exception of strain JCM 3034 (Fig. 1a), but not in other genera of the family (Table 1). Interestingly enough, Pseudonocardia, the only genus of the former PSF (Pseudonocardia, Saccharopolyspora, Faenia) group (Embley, 1988a), lacked this glycolipid. The results are in good agreement with the reclassification of Faenia rectivirgula as Saccharopolyspora rectivirgula (Korn-Wendisch et al., 1989) leaving this genus and Pseudonocardia as distinct (though closely related) taxa of actinomycetes (Embley et al., 1988b; Embley, 1992). The major glycolipid occurring in the cells of Saccharopolyspora seemed therefore valuable for the differentiation of the actinomycetes from allied taxa. For this reason the glycolipid marker from the type strain of the type species Saccharopolyspora hirsuta was chosen for the preliminary chemical studies. Preliminary chemical characterisation of the glycolipid. The major glycolipid from Saccharopolyspora hirsuta ATCC 27875, Rf value 0.6 (Fig. 1b), was isolated and purified. Figure 1. TLC of glycolipids from some saccharopolysporae: (a) 1-4: Saccharopolyspora rectivirgula, strains JCM 3057, JCM 3099, A164 and JCM 3034 (the only strain lacking the major glycolipid (G); (b) Saccharopolyspora hirsuta ATCC 27875 (formerly K52) used for isolation of the major glycolipid (G). Solvent system: chloroform-methanol-water (65:25:4, v/v). Detection: orcinol spray (0.5% orcinol in ethanol with 3% H2SO4) followed by heating at 120 C. Its purity was confirmed by two-dimensional TLC in two pairs of solvents (Fig. 2). After hydrolysis of the glycolipid its sugar analysis revealed the presence of mannose and glycerol in the molar ratio 2:1, respectively. The total amount of neutral sugars of the glycolipid was 22%. The results of the quantitative analyses of fatty acids (%) were as follows:
Branched fatty acids
anteiso C17:0 49.44
iso C17:0 31.22
iso C16:0 12.99
Normal fatty acids (as minor components)
C16:0 2.91
C17:0 1.23
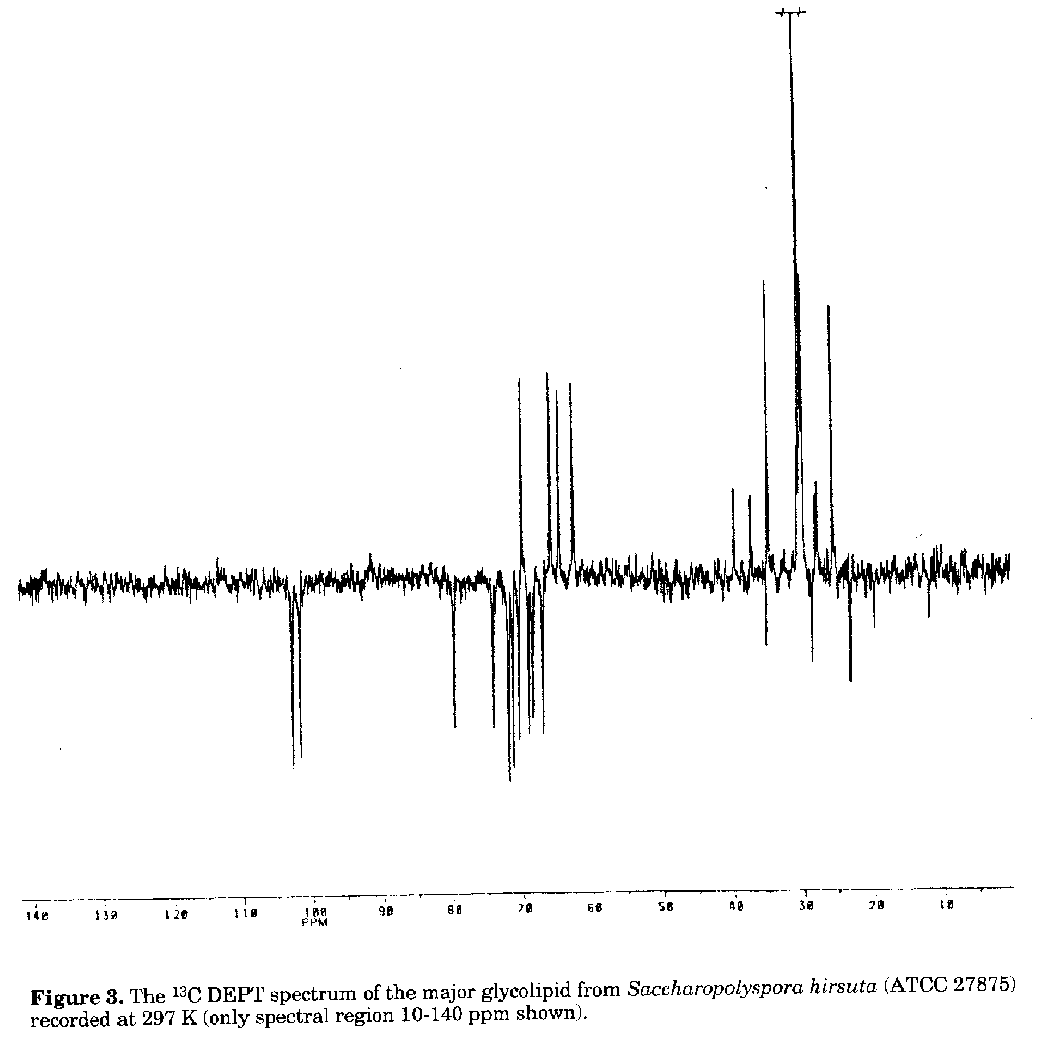
C18:0 2.21Figure 2. Two-dimensional TLC of the purified major glycolipid (G) from Saccharopolyspora hirsuta (ATCC 27875). Solvent systems: first direction, chloroform-methanol- water (65:25:4, v/v); second direction, chloroform-acetic acid-methanol-water (80:15:12:4, v/v); vanillin as spray reagent (0.5% vanillin in 3% ethanol with H2SO4) followed by heating 120 C. The mono-methyl branched fatty acids of the iso- and anteiso- series were in agreement with the major compounds of fatty acid spectrum from whole cells of Saccharopolyspora hirsuta (Embley et al., 1987) and of other species of the genus (Goodfellow et al., 1989; Korn-Wendisch et al., 1989). The glycolipid was subjected to mild alkaline hydrolysis to isolate the sugar moiety. Complete de-O-acylation was achieved after three days and the result was monitored by the appearance of oligosaccharide and the disappearance of glycolipid spots on TLC silica gel plates. When the oligosaccharide moiety of the compound was treated with a-mannosidase, free mannose and glycerol were obtained, showing that mannose residues had an a-anomeric configuration. Methylation of the oligosaccharide resulted in two derivatives in equal proportions, namely 2,3,4,6-tetra-O- methyl-1,5-di-O-acetyl mannitol and 2,4,6-tri-O-methyl-1,3,5- tri-O-acetyl mannitol. This indicated that one terminal and one 3-substituted mannose were present. The enzymatic procedure used to determine the absolute configuration of mannose, based on its phosphorylation by hexokinase, revealed that the sugar present in the glycolipid hydrolysate had a D- configuration. To sum up, the sugar moiety of the glycolipid appeared to be disaccharide with the formula: alpha -D Manp (1->3)-alpha-D-Manp (1- >3) Gro where Man and Gro stand for mannose and glycerol, respectively. The 13C-NMR spectrum recorded in DEPT mode (Doddrell et al., 1982) proved the a-anomery of mannose residues, since the chemical shift for C-1 of terminal mannose was at 102.6 ppm and that of 3-linked mannose was at 101.5 ppm (Fig. 3). These results are in agreement with literature data (Bock et al., 1984). Initial investigations using fast atom bombardment-mass spectrometry (FAB-MS) indicated that the internal mannose in the glycolipid was substituted by a fatty acid residue (Gamian et al., 1992). The uncommon structure, where one of the fatty acids is attached to the sugar residue, was also found in some diacylglycerides isolated from a strain of Streptomyces (Batrakov et al., 1978). Further studies are in progress to determine the more detailed structure of the major glycolipid. A comparison with compounds of other Saccharopolyspora species seems also to be necessary. Figure 3. The ^13C DEPT spectrum of the major glycolipid from Saccharopolyspora hirsuta (ATCC 27875) recorded at 297 K (only spectral region 10-140 ppm shown). ACKNOWLEDGEMENTS This work was partially supported by Grant (No 4 0646 91 01) from the Committee for Research and Science (KBN), Poland. REFERENCES Batrakov, S.G. & L.D.Bergelson (1978). Lipids of streptomycetes. Structural investigation and biological interrelation. Chem.Phys.Lipids, 21: 1-29 Bock, K., C.Pedersen & H.Pedersen (1984). Carbon-13 nuclear magnetic resonance data for oligosaccharides. Adv.Carbohydr.Chem.Biochem., 42: 193-225 Doddrell, D.M., D.T.Peg & M.R.Bendal (1982). Distortionless enhancement of NMR signals by polarization transfer. J.Magn.Reson., 48: 323-327 Embley, T.M. (1992). The family Pseudonocardiaceae. In: A.Balows, H.G.Trper, M.Dworkin, W.Harder & K.H.Schleifer (eds.) The Prokaryotes. Springer Verlag, New York, Vol.1, pp. 996-1027 Embley, T.M., A.G.O'Donnel, J.Rostron & M. Goodfellow (1988a). Chemotaxonomy of wall type IV actinomycetes which lack mycolic acids. J.gen.Microbiol., 134: 953- 960 Embley, T.M., J.Smida & E.Stackebrandt (1988b). Reverse transcriptase sequencing of 16S ribosomal DNA from Faenia rectivirgula, Pseudonocardia thermophila and Saccharopolyspora hirsuta, three wall type IV actinomycetes which lack mycolic acids. J.gen.Microbiol., 134: 961-966 Embley, T.M., R.Wait, G.Dobson & M.Goodfellow (1987). Fatty acid composition in the classification of Saccharopolyspora hirsuta. FEMS Microbiol.Lett., 41: 131-135 Gamian, A., H.Mordarska, J.Ulrich & J.Defaye (1992). Structural studies of the major glycolipid of Saccharopolyspora hirsuta. Abstr.XVI Int.Carbohydrate Symp., Paris, July 5-10, C143, p. 608 Gamian, A., E.Romanowska, U.Dabrowski & J.Dabrowski (1991). Structure of the O-specific, sialic acid containing polysaccharide chain and its linkage to the core region in lipopolysaccharide from Hafnia alvei strain 2 as elucidated by chemical methods, gas-liquid chromatography/mass spectrometry and 1H NMR spectroscopy. Biochemistry, 30: 5032-5038 Gamian, A., E.Romanowska, U.Dabrowski & J.Dabrowski, J. (1993). Structure of the O-specific polysaccharide containing pentitol phosphate, isolated from Hafnia alvei strain PCM 1191 lipopolysaccharide. Eur.J.Biochem., 213: 1255-1260 Goodfellow, M., J.Lacey, M.Athalye, T.M. Embley & T.Bowen (1989). Saccharopolyspora gregorii and Saccharopolyspora hordei: two new actinomycetes species from fodder. J.gen.Microbiol., 135: 2125-2139 Kates, M. (1986). Techniques of lipidology: isolation, analysis and identification of lipids. In: R.H.Burdon & P.H. van Knippenberg (eds.) Laboratory Techniques in Biochemistry and Molecular Biology. Elsevier, Amsterdam, Vol. 3, part 2, pp. 100-112 Korn-Wendisch, F., A.Kempf, E.Grund, R.M. Kroppenstedt & H.J.Kutzner (1989). Transfer of Faenia rectivirgula Kurup and Agre 1983 to the genus Saccharopolyspora Lacey & Goodfellow 1975, elevation of Saccharopolyspora hirsuta subsp.taberi Labeda 1987 to species level and emended description of the genus Saccharopolyspora. Int.J.Syst.Bacteriol., 39: 430-441 Mordarska, H. (1985/1986). Taxonomic value of glycolipids of actinomycetes and allied taxa. The Actinomycetes, 19: 11-32 Mordarska, H., A.Gamian & I.Ekiel (1992). Structural studies of major glycolipids from Curtobacterium flaccumfaciens pathovar betae and Rothia dentocariosa. Actinomycetes, 3: 31-36.
Copyright 1995 CETA
The following images related to this document are available:Halftone images[ac95001b.gif] [ac95001a.gif]Photo images[ac95001b.jpg] [ac95001a.jpg]Line drawing images[ac95001c.gif] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}