 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
PARTIAL CHARACTERIZATION OF STREPTOMYCES PHAGES ISOLATED FROM THE SOILS OF JARRAH FOREST IN WESTERN AUSTRALIA K.A. EL-TARABILY, D.I. KURTBOKE and G.E.ST.J. HARDY Murdoch University, School of Biological and Environmental Sciences, Perth, W.A. 6150, Australia
Code Number: AC95002
Size of Files:
Text: 29K
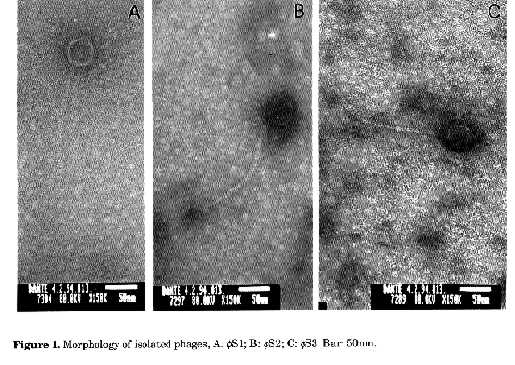
Graphics: Photos (jpg) -60.7K / Halftones (gif) -83KNOTE: Strains designated 'f' below are designated 'theta' in the printed edition. ABSTRACT. Three phages fS1, fS2 and fS3 which utilized Streptomyces diastaticus, S.griseus and S.hygroscopicus respectively, as propagation hosts, were isolated from the soils of jarrah forest in Western Australia. They were partially characterized through their physiochemical properties, plaque morphology, host range and particle morphology. Host range of these three Siphoviridae (B1) morphotype phages demonstrated a wide activity spectra within the genus Streptomyces. Adsorption rate constants and burst sizes of the three phages were in the range of 1.58x10-7, 1.26x10-7 and 5.97x10-9 ml/min and 15.5, 22.5, 12.9 virions/cell, respectively. Agarose electrophoresis of restriction endonuclease digests of the phage DNAs indicated that the three phages were different. Diaz et al. (1989) indicated that the isolation and identification of Streptomyces phages are of interest for a variety of reasons, which include (i) the problems they cause to fermentation industries (Chater, 1986), (ii) their value for typing streptomycetes in taxonomic studies (Wellington and Williams, 1981; Prauser, 1984; Korn-Wendish and Schneider, 1992), (iii) their use for the detection and understanding of host controlled restriction-modification systems (Diaz et al., 1989), (iv) their utilization as tools for genetic exchange and analysis in Streptomyces (Herron and Wellington, 1990), (v) the study of their general and molecular biology (Lomovskaya et al., 1980) and ecology (Williams et al., 1987) and most recently (vi) their use to reduce the numbers of streptomycetes on isolation plates to select rare actinomycetes (Kurtb”ke et al., 1992). In an attempt to isolate actinoplanetes from jarrah forest soils of Western Australia, we used three polyvalent streptomycete phages to reduce the numbers of streptomycetes on isolation plates to select the targeted taxon. This paper deals with the partial characterization of these phages and describes their host range and physiochemical properties. MATERIALS AND METHODS Soil samples. Fresh soil samples were collected from the A horizon of the northern jarrah (Eucalyptus marginata Donn ex Sm.) forest (near Dwellingup) in Western Australia. These samples were mixed to form a bulk sample with a final pH of 6.5 and this heterogeneous mixture was used to isolate phages. Propagation hosts. Type strains of Streptomyces diastaticus (ATCC 3315), S. griseus (NCTC 7807) and S.hygroscopicus (ATCC 31955) were obtained from Dr.E. Lacey, McMaster Laboratory, C.S.I.R.O., Sydney. Cultures were maintained on oatmeal agar + 0.1% yeast extract (BBL, Becton Dickinson, USA) (Williams and Wellington, 1982). Inocula were stored as spore suspensions in 10% glycerol at -20 C (Wellington and Williams, 1978). Phage isolation and purification. Flasks (250ml) containing 20ml of sterile peptone-yeast extract calcium (PYCa) broth (Bradley et al., 1961) were inoculated with 1ml of the spore suspension of the prospective host streptomycete and 2g of the bulk soil sample. These were then incubated in a gyrotory shaker (Model G76, New Brunswick Scientific-Edison, N.J., USA) at 200rpm for 2dd at 28 C. After incubation, the suspensions from each flask were centrifuged for 1hr at 2,000g and the supernatants filtered through sterile Millipore membranes of pore size 0.22æm (Millipore Corp.) and collected in sterile tubes. Filtrates (0.2ml) were spotted onto plates of PYCa agar (Bradley et al., 1961) on which 0.3ml of glycerol suspension of the prospective host had been spread onto PYCa plates and dried for 30min in a laminar flow (Vickers and Williams, 1987). The plates were then incubated at 28 C for 2dd and examined for plaques (Williams et al., 1980). Single plaques were removed from the agar plates and resuspended in 1 ml of PYCa broth for 36hrs (Williams et al., 1980). A sample of this broth was then filtered and spotted on the PYCa plates previously inoculated with the prospective hosts. Single plaques which developed on these plates were removed, resuspended, filtered and these purified phage suspensions were stored at 4 C (Williams et al., 1980). Plaque morphology. Phage suspensions (0.2 ml) which contained 107 plaque forming units (pfu/ml) were spotted onto PYCa agar seeded with the host Streptomyces species suspensions (x106 cfu/ml). The plates were then incubated for 48hrs at 28 C and plaque morphology was examined. Host range. Type strains of Streptomyces spp. used were obtained from Dr.E. Lacey, McMaster Laboratory, C.S.I.R.O., Sydney (Table 1). The host range of the phages were studied by spotting 0.2ml of phage suspensions containing 107 pfu/ml onto PYCa agar plates each previously seeded with a glycerol suspension (x106 cfu/ml) of one of the type strains (Table 2). Phage suspensions were prepared by ten-fold dilutions of the clear spots on the propagation host. Clear spot dilution (CSD) is equivalent to the routine test dilution (RTD) (Wellington and Williams, 1981). The plates were then incubated for 2dd at 28 C and examined for lysis. Electron microscopy. A drop of each phage suspension (107 pfu/ml) was placed on 200-mesh copper grids with carbon-coated Formvar films and the excess drawn off with filter paper. A saturated solution of uranyl acetate was then placed on the grids and the excess drawn off as before. Specimens were observed with a JEOL-2000 FX II transmission electron microscope operated at 80 kV. Effects of physical and chemical agents on phage propagation and viability. Three different media: peptone- yeast extract (Bradley et al., 1961), Nutrient agar (NA) (Difco) and Tryptic soy agar (TSA) (Difco) supplemented with and without Ca(NO3)2 (0.05%) and with two different concentrations of NaCl (0.1M, 0.01M) (Brownell et al., 1967) were tested to determine the effects of these complex media on phage propagation. The effects of physical and chemical agents on the phage and the effects of the size of phage inoculum and host age on the number of plaque forming units were tested according to the methods described by Brownell et al. (1967). Adsorption and one-step growth experiment. The adsorption rates of the three phages were determined by measuring residual plaque-forming ability in membrane-filtered samples of an attachment mixture (Dowding, 1973) and the adsorption rate constant K ml/min was calculated (Sykes et al., 1981). A one-step growth experiment was conducted as described by Dowding (1973). Restriction endonuclease digestion and agarose gel electrophoresis. The nucleic acids of the three phages were isolated according to the methods described by Rodriguez and Tait (1983). The isolated nucleic acids and lDNA were digested with the enzymes Hind III, Bam HI (Gibco BRL) and Eco RI (Progen Industries Ltd.) at 37 C for 3hrs (Rodriguez and Tait, 1983). Reaction buffers were supplied with the enzymes. The intact and enzyme treated nucleic acids were electrophoresed in 1% agarose (Progen Industries Ltd.) in 40 mM Tris-acetate, pH 8.0 and 2mM EDTA. Electrophoresis was run on a horizontal apparatus at 8V/cm for 2hrs at room temperature (Rodriguez and Tait, 1983). After electrophoresis, the gel was stained with ethidium bromide (0.5æg/ml) and photographed. lDNA was used as size marker during the examination of the restriction endonuclease digested phage DNAs. RESULTS Plaque morphology. The phages fS1 specific to S.diastaticus (ATCC 3315) and fS2 specific to S.griseus (NCTC 7807) formed small circular clear plaques (0.8 and 0.7 mm respectively). Phage fS3, specific to S.hygroscopicus (ATCC 31955), formed round plaques with sharp edges 1mm in diameter. Electron microscopy. Negatively stained particles of the three phages fS1, fS2 and fS3 belonged to the Siphoviridae (B1) morphotype (Francki et al., 1991) and the phages fS1, fS2 had icosahedral heads 41.2x41.2 and 41.2x41.2nm in diameter respectively. However, fS3 had an ovoid head (47.0x32.3nm). The tails of the phages fS1, fS2 and fS3 were 7.0x 159, 7.0x188 and 5.9x253nm respectively (Figs. 1, A, B and C).
SPECIES STRAIN fS1 fS2 fS3
Streptomyces albaduncus CBS 698.72 - - -
Streptomyces albidoflavus CBS 416.34 + - -
Streptomyces albidopureus CBS 353.79 + + +
Streptomyces albocolor CBS 354.79 - - -
Streptomyces albus ATCC 3004 + - -
Streptomyces avermitilis ATCC 31267 + + +
Streptomyces bambergiensis CBS 780.72 + + +
Streptomyces diastaticus ATCC 3315 PH + +
Streptomyces erumpens CBS 252.65 + + +
Streptomyces flaveolus ATCC 3319 + + +
Streptomyces griseus NCTC 7807 + PH +
Streptomyces hygroscopicus ATCC 31955 + + PH
Streptomyces lisandri ATCC 27963 + + +
Streptomyces pyridomyceticusCBS 936.68 - - +
Streptomyces sp. ATCC 31587 + + +
Streptomyces torulosus ATCC 29340 + + +
Streptomyces vastus CBS 648.69 + + +
Streptoverticillium netropsis
ATCC 23940 - - -
Streptoverticillium rectiverticillatum
CBS 951.69 - - -
Streptoverticillium roseoverticillatum
CBS 648.69 - - -
Table 1. Host range of Streptomyces phages
isolated from the soils of jarrah forest (+: host species
susceptible to phage lysis; -: host species not susceptible to
phage lysis; PH: propagation host; CBS: Centraalbureau voor
Schimmelcultures, Baarn, The Netherlands; ATCC: American Type
Culture Collection, Rockville, Maryland; NCTC: National
Collection of Type Cultures, London).Host range. Phages fS1, fS2 and fS3 were not species specific and lysed a wide range of Streptomyces species. However, they did not utilize Streptoverticillium species (Table 1). Effect of physical and chemical agents on phage propagation and viability. PYCa supplemented with 0.05% Ca(NO3)2 was the best medium for phage propagation followed by NA and TSA (Table 2). Increased NaCl concentrations reduced the numbers of congruent plaques on all three media and no plaque forming units were counted on TSA when it was supplemented with 0.1M NaCl (Table 2). The three phages were sensitive to chloroform, thymol, hydrogen peroxide and ethanol, all of which greatly reduced the numbers of plaque forming units (Table 2). Freezing at -4 C for 2hrs had an adverse effect on phage titre in particular on phage fS2. At 4 C phage numbers decreased markedly with time. Less phages were obtained when propagation hosts were incubated for 0-5hrs, in comparison to those incubated for 10-12hrs (Table 3). Greater levels of host inoculum resulted in greater phage titres and similarly greater phage inoculum size resulted in greater phage output (Table 3). Adsorption rate constant, latent period and burst size of the phages. Adsorption rates of the three phages rising from spores were 1.58x10^-7, 1.26x10^-7 and 5.97x10^-9 ml/min respectively. The percentage decrease observed in free phage numbers was linear between 0 and 20 min. After 20min the adsorption rates appeared to fall. Counts from samples taken after 35-40min suggested that infected cells were lysing and liberating phage. Latent period values, obtained for the three phages, were 35, 40 and 40min, which confirmed the observations made during the adsorption experiments.
-----------------------------------------------------------
MEDIUM SUPPLEMENT pfu/ml of phage suspension
fS1 fS2 fS3
----------------------------------------------------------
Peptone-yeast extract
0.1 M NaCl - - 1.0x10^2
0.01 M NaCl 4.5x10^3 3.0x10^4 2.0x10^4
0.05% (w/v) Ca(N03)2 2.0x10^6 1.8x10^7 7.7x10^7
0.1 M NaCl 7.5x10^2 - -
Nutrient agar
0.01 M NaCl 2.5x10^4 6.0x10^3 2.5x10^4
0.05% (w/v) Ca(N03)2 12.5x10^4 9.0x10^5 4.0x10^5
Tryptic soy agar
0.1 M NaCl - - -
0.01 M NaCl 5.0x10^3 - -
0.05% (w/v) Ca(N03)2 3.6x10^4 - -
TREATMENT
Untreated control 2.0x10^6 1.8x10^7 7.7x10^7
Chloroform (biphasic: 1 vol. chloroform to 2 vol. water)
2.8x10^4 4.0x10^2 8.5x10^3
Thymol (0.l g in l0ml water) 5.0x10^2 1.1x10^1 1.7x10^3
Hydrogen peroxide (3% v/v, in water) - - -
Hydrogen peroxide (0.3% v/v, in water)
1.5x10^2 - -
Ethanol (30% v/v, in water) 8.5x10^3 1.2x10^1 1.4x10^3
Freezing (2hrs) 1.2x10^4 1.5x10^1 8.0x10^4
4 C for 1 d 2.0x10^6 1.8x10^7 7.7x10^7
4 C for 7 dd 1.4x10^5 2.3x10^6 3.1x10^5
4 C for 14 dd 2.3x10^3 3.5x10^4 2.4x10^3
4 C for 30 dd 1.2x10^1 1.3x10^1 1.9x10^1
45 C for 15 min 6.0x10^5 1.4x10^6 1.2x10^6
56 C for 15 min 1.8x10^3 1.3x10^3 2.4x10^4
Ultrasonic treatment (20kc/sec) for 60 sec
1.3x10^6 1.5x10^6 6.4x10^7
UV irradiation for 30sec 1.8x10^6 1.1x10^7 6.8x10^7
UV irradiation for 90sec 5.0x10^4 1.0x10^5 1.5x10^5
UV irradiation for 150sec 8.0x10^2 1.7x10^2 1.8x10^3
UV irradiation for 210sec - - -
--------------------------------------------------------------
Table 2. Effects of complex media, chemical and
physical agents on the viability and propagation of
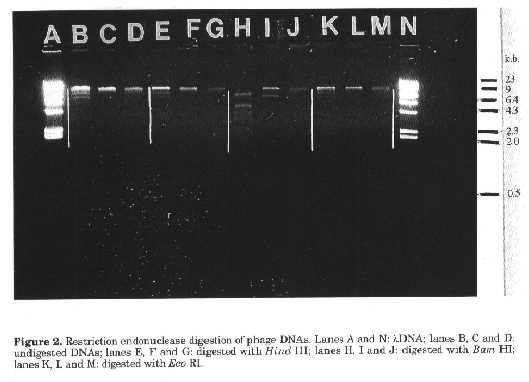
phage ( - :no plaque forming units counted).The rise periods of the phages were 40, 30 and 40min respectively, with average burst sizes of 15.5, 22.5 and 12.9 virions/cell. The second burst began at around 70-80min, nearly 30-40min after the first burst began, which confirmed the minimum latent period of 30-40min for the three phages (Table 4). Restriction endonuclease digestion of phage DNAs. The phages f S1, fS2 and fS3 were readily digested by Bam HI but not the other enzymes used (Fig. 2). The three phages did not share similar restriction fragments indicating that they were different. This was supported by particle morphology and the results obtained in growth kinetics experiments (Table 4). DISCUSSION Phages active against Streptomyces spp., the predominant actinomycete genus in soil, are readily detected and it appears that streptomycete phages are widespread in the soil environment (Williams and Lanning, 1984).
Incubation time (hrs) of host pfu/ml of phage suspension
prior to inoculation fS1 fS2 fS3
0 1.4x10^6 1.9x10^7 6.1x10^7
3 1.3x10^6 1.6x10^7 8.2x10^7
5 3.2x10^6 2.8x10^7 9.5x10^7
10 2.4x10^8 1.9x10^9 1.4x10^9
12 3.5x10^8 2.5x10^9 2.8x10^9
--------------------------------------------------------------
Propagation host inoculum size
(cfu/ml of host spore suspension)
10^7 1.8x10^6 1.2x10^7 6.4x10^7
10^6 2.6x10^5 2.7x10^6 3.2x10^6
10^5 3.1x10^3 3.6x10^3 2.5x10^4
10^4 1.4x10^2 1.5x10^1 1.3x10^2
--------------------------------------------------------------
Phage inoculum size
(pfu/ml of phage suspension)
10^7 1.3x10^6 1.1x10^7 5.2x10^7
10^6 1.0x10^6 1.0x10^7 3.1x10^7
10^5 2.4x10^5 2.4x10^6 1.4x10^6
10^4 2.6x10^3 1.3x10^5 2.8x10^5
10^3 1.1x10^3 2.4x10^3 3.1x10^3
10^2 2.7x10^2 1.8x10^2 1.9x10^2
10^1 1.6x10^1 1.5x10^1 1.9x10^1
Table 3. Effects of age and inoculum size of
propagation hosts on phage production and effects of phage
inoculum size on the yield of phages. Propagation hosts used
for phages fS1, fS2 and fS3 were Streptomyces diastaticus
(ATCC 3315),S.griseus (NCTC 7807) and
S.hygroscopicus (ATCC 31955) respectively.-----------------------------------------------------------
Phage Adsorption rate Latent period Rise period Burst size
constant K (min) (min) relative
(ml.min) increase
(virions/cell)
-----------------------------------------------------------
fS1 1.58x10^-7 35 40 15.5
fS2 1.26x10^-7 40 30 22.5
fS3 5.97x10^-9 40 40 12.9
Table 4. Biological properties of Streptomyces
phages.------------------------------------------------------------ The rapid isolation of Streptomyces phages without any enrichment techniques agrees with these previous findings. The isolated phages are of the same morphological type as other streptomycete phages (Ackermann et al., 1985). The broad activity spectra are in agreement with the previous findings that common phage susceptibility is one of the distinctive characters of the members of the genus Streptomyces (Wellington and Williams, 1981; Prauser, 1984).
The burst sizes obtained for the phages were lower than the ones obtained by Dowding (1973) but higher than some of the values obtained by Sykes et al. (1981) for phages that utilized neutrophilic hosts. However, as observed by Jones and Bradley (1965), many factors can influence phage burst size, such as the filamentous nature of the host, spore clumping and host age. Dowding (1973) also concluded that actinomycetes, including representatives of the genus Streptomyces, do not germinate synchronously, with the result that by the time a high enough proportion of spores have germinated, some of them will have formed large clumps of mycelium. These clumps can adsorb a great many phage particles yet continue to plate out as single colony-forming units. Therefore, the concept of multiplicity of infection will remain meaningless in actinomycete host-phage systems until a method is found to achieve synchronous and uniform spore germination. This is the first record of the detection of Streptomyces phages from the jarrah forest soils of Western Australia. As indicated by Williams and Lanning (1984) phages are a neglected ecological entity. Isolation of phages from different substrates in neglected environments will increase our knowledge on the ecology of species, genus and family specific phages and provide additional information on phage ecology and phage-host interactions (Williams and Lanning, 1984; Kurtb”ke and Williams, 1991). ACKNOWLEDGEMENTS. This work was sponsored by Alcoa Australia Ltd. We thank Dr. I.Colquhoun for his encouragement. Mr.S.Wylie for his kind help during the restriction endonuclease digestion of the phage DNAs and Prof. M.J.K.Jones for allowing us to use his facilities. Mr.S.Parry (University of Western Australia, Electron Microscopy and Microanalysis Centre) assisted with the preparation of the electron micrographs. REFERENCES Ackermann, H.-W., L.Berthiaume & L.A. Jones (1985). New actinophage species. Intervirol., 23: 121-130 Adams, M. (1959). Bacteriophages. Wiley InterScience, New York Bradley, S.G., D.L.Anderson & L.A.Jones (1961). Phylogeny of actinomycetes as revealed by susceptibility to actinophage. Dev.Ind.Microbiol., 2: 223-237 Brownell, G.H., J.N.Adams & S.G.Bradley (1967). Growth and characterization of nocardiophages for Nocardia canicruria and Nocardia erythropolis mating types. J.gen.Microbiol., 47: 247-256 Chater, K.F. (1986). Streptomyces phages and their applications to Streptomyces genetics. In: S.W.Queener & L.E.Day (eds.) The Bacteria: A Treatise on Structure and Function. Vol.IX: Antibiotic Producing Streptomyces. Academic Press, New York, pp. 119-158 Diaz, L.A., C.Hardisson & M.R.Rodicio (1989). Isolation and characterization of actinophages infecting Streptomyces species and their interaction with host restriction-modification systems. J.gen.Microbiol., 135: 1847-1856 Dowding, J.E. (1973). Characterization of bacteriophage virulent for Streptomyces coelicolor A3(2). J.gen.Microbiol., 76: 163-176 Francki, R.I.B., C.M.Fauquet, D.L.Knudson & F.Brown (1991). Classification and nomenclature of viruses. Fifth report of the International Committee on Taxonomy of Viruses. Arch.Virol., Suppl. 2: 161-166 Gold, W. (1959). Effects of the medium and its composition on the activities of actinophage for Streptomyces griseus. Ann.N.Y.Acad.Sci., 81: 995-1016 Goyal, S.M. (1987). Methods in phage ecology. In: S.M.Goyal, C.P.Greba & G.Bitton (eds.) Phage Ecology. John Wiley & Sons, Inc., New York, pp. 267-288 Herron, P.R. & E.M.H.Wellington (1990). New method for extraction of streptomycete spores from soil and application to the study of lysogeny in sterile amended and nonsterile soil. Appl. Environ.Microbiol., 56: 1406-1412 Jones, L.A. & S.G.Bradley (1965). The life-cycle of an actinophage for Streptomyces venezuelae. J. gen.Microbiol., 40: 191-198 Korn-Wendish, F. & J.Schneider (1992). Phage typing - a useful tool in actinomycete systematics. Gene, 115: 243-247 Kurtb”ke, D.I. & S.T.Williams (1991). Use of polyvalent phage for selective isolation purposes - current problems. Actinomycetes, 2: 31-34 Kurtb”ke, D.I., C.-F.Chen & S.T.Williams (1992). Use of polyvalent phage for reduction of streptomycetes on soil dilution plates. J.App. Bacteriol., 72: 103-111 Lomovskaya, N.D., K.F.Chater & N.M.Mkrtumian (1980). Genetics and molecular biology of Streptomyces bacteriophages. Microbiol.Revs., 44: 206-229 Prauser, H. (1984). Phage host ranges in the classification of Gram-positive branched and related bacteria. In: L.Ortiz- Ortiz, J.F.Bojalil & V.Yakoleff (eds.) Biological, Biochemical and Biomedical aspects of Actinomycetes. Academic Press, Orlando, pp. 617-633 Rodriguez, R.L. & R.C.Tait (1983). Recombinant DNA Techniques: An Introduction. The Benjamin/Cummings Publ.Co., Inc., California Reading, Mass., USA, pp. 53-77 Sykes, I.K., S.Lanning & S.T.Williams (1981). The effect of pH on soil actinophage. J.gen.Microbiol., 122: 271-280 Vickers, J.C. & S.T.Williams (1987). An assessment of plate inoculation procedures for the enumeration and isolation of soil streptomycetes. Microbios Lett., 35: 113-117 Walton, R.B. (1951). Effects of cations upon multiplication of actinophage for Streptomyces griseus. Antibiot.Chemother., 1: 518-522 Wellington, E.M.H. & S.T.Williams (1978). Preservation of actinomycete inoculum in frozen glycerol. Microbios Lett., 6: 151-157 Wellington, E.M.H. & S.T.Williams (1981). Host ranges of phages isolated to Streptomyces and other genera. Zentralbl.Bakteriol.Mikrobiol. Hyg. I.Abt. Suppl., 11: 93-98 Williams, S.T. & S.Lanning (1984). Studies on the ecology of streptomycete phage in soil. In: L. Ortiz-Ortiz, J.F.Bojalil & V.Yakoleff (eds.) Biological, Biochemical and Biomedical Aspects of Actinomycetes. Academic Press, Orlando, pp. 473-483 Williams, S.T., A.M.Mortimer & L. Manchester (1987). Ecology of soil bacteriophages. In: S.M.Goyal, C.P.Greba & G.Bitton (eds.)Phage Ecology. John Wiley & Sons, Inc., New York, pp. 157-179 Williams, ST., E.M.H.Wellington & L.S.Tipler (1980). The taxonomic implications of the reactions of representative Nocardia strains to actinophage. J.gen.Microbiol., 119: 173-178 Williams, S.T. & E.M.H.Wellington (1982). Actinomycetes. In: A.Page, R.H.Miller & O.R.Keency (eds.) Methods of Soil Analysis. Part 2: Chemical and Microbiological Properties. Am.Soc. Agronomy & Soil Sci.Soc. of America, Madison, 2nd ed., pp. 969-987. Copyright 1995 CECT
The following images related to this document are available:Halftone images[ac95002b.gif] [ac95002a.gif]Photo images[ac95002a.jpg] [ac95002b.jpg] |
| |||||||||
{kind=link}
{kind=link}