 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
MUTAGEN - INDUCED CHANGES IN CELL COMPONENTS AND BIOSYNTHESIS OF ANTIBIOTIC 1012 IN STREPTOMYCES SPECTABILIS
V. IVANOVA, R. ROUSEVA, M. NAIDENOVA, R. RACHEV and M. KOLAROVA
Institute of Microbiology, Bulgarian Academy of Sciences, 1113 Sofia, Bulgaria
ABSTRACT. Twelve actinomycete strains (parent organisms, mutants and a fusant) of Streptomyces spectabilis were investigated with reference to the biosynthesis of antibiotic complex 1012. Mutant H-72, obtained by UV radiation, exhibited the highest productivity of the antibiotic complex. Correlation between production of complex 1012 and biosynthesis of cell structural components (lipids, polysaccharides and proteins) was investigated.
Strains of Streptomyces spectabilis produce a variety of antibiotics active against Gram-positive and Gram-negative bacteria. Strain NRRL 2494, forms streptovaricin, spectinomycin and prodigiosin (Gauze et al., 1983), strain B 44 streptovaricin (Yamazaki, 1968a, 1968b) and strain NRRL 2792 spectinomycin (Mason et al., 1961). Derivatives of aureolic acid, known as spectomycin, and cyclic depsipeptides (deoxymonomycins) have been isolated from a mutant strain of the same species (Staley, 1990). Preliminary investigations have shown that S.spectabilis strains 1000 and 1011 produce the antibiotic complex 1011 (identical to the streptovaricin complex), prodigiosin and the antibiotic complex 1012 (Ivanova et al., 1987, 1989, 1991). The aim of the present study was to investigate the possible correlation between cell composition (lipids, polysaccharides and proteins) and production of antibiotic complex 1012 in parental strains of S.spectabilis and in their mutants. MATERIALS and METHODS Organisms and growth conditions. Two S.spectabilis strains were investigated: 1000 (H-I), obtained from the Collection of the Institute of Microbiology of the Bulgarian Academy of Sciences, and 2494 (H-II-3-9) from the National Collection of Industrial Microorganisms and Cell Cultures, Bulgaria. Mutants H-1, H-2, H-3, H-4, H-13 and H-14 were obtained from strain 1000 by treatment with 10 (strain H-14), 50 (strains H-1 and H-4), 70 (strain H-13) and 100g/ml (strains H-2 and H-3) of streptovaricin on Bennett agar (Dietz et al., 1963). Mutants H-72, H-33 and H-29 were obtained by UV-radiation (0.44, 0.33 and 0.22 x 10^-5 J/mm^2 respectively) of a 7-day spore suspension of strain 1000. Protoplast fusion (fusant H-f) was carried out between tyrosine and streptomycin-resistant auxotrophs of strain 1000 and a histidine, pyridoxine and streptomycin sensitive mutant of strain 2494. Tyrosine and histidine were used as markers. Protoplast regeneration was obtained in R2YE medium (Thompson et al., 1982). All strains were grown in Bennett liquid medium using a rotary shaker at 220 r.p.m. at 28-30 C for 96hrs. Antibiotic complex 1012. Mycelia were extracted three times with 50ml methanol. Combined extracts were concentrated to dryness at 45 C under reduced pressure and dissolved in methanol (4ml), filtered and precipitated with acetone (1:5 v/v). Precipitates were allowed to stand for 24hrs at 5-10 C, filtered, washed with acetone, dissolved in 2ml methanol and analysed by thin-layer chromatography (TLC). TLC of the antibiotic complex was performed on silica gel (G60, F[254], Merck) with ethyl-methyl ketone/ethanol/ammonia (1:1:1). Spots were visualised with concentrated H2SO4 and heating at 120 C for 5-8min. Lipids. Lipids were isolated by the Blight-Dyer method (Christie, 1973). Fatty acid methyl esters were obtained by transesterification with BF3-CH3OH according to the Metcalf method (Christie, 1973). Fatty acid methyl esters were analysed with a Carlo Erba Science 4300 apparatus equipped with a fused silica capillary column. Operating conditions of the instrument were as follows: initial temperature 90 C (5 min), programme rate 4 C/min, final temperature 220 C (10min), N2 (4ml/min), detection by flame ionisation. Polysaccharides. Mycelia were dried at 60 C and extracted three times with 0.5M NaOH for 3hrs. The combined extracts were precipitated with absolute ethanol (1:2 v/v), stored for 24hrs at 5-10 C, centrifuged and the sediment dissolved in 8ml 0.5M NaOH. After precipitation with absolute ethanol (1:1 v/v) and storage for 48hrs at 5-10 C, the precipitate was centrifuged, dissolved in 5ml 0.5M NaOH and the solution treated with 15% (w/v) trichloracetic acid (1:2 v/v) for 4hrs. Protein containing precipitates were removed by centrifugation, the supernatant precipitated with acetone (1:2 v/v) and the resulting precipitate centrifuged, dissolved in distilled water and dialysed by running water (72hrs) and distilled water (168hrs). Aqueous solutions were lyophilised. The purity of the isolated polysaccharides with reference to the presence of proteins was checked after acid hydrolysis with 6M HCl. The absence of amino acids was established by TLC. For the analysis of carbohydrates present in the polysaccharides, samples (35mg) were hydrolysed in 2M H2SO4 (7ml) for 8hrs at room temperature and heated at 110 C for 1hr. Reaction mixtures were neutralised with saturated Ba(OH)2 solution to pH 7.0. BaSO4 precipitates were filtered and washed with water. The filtrates were concentrated under vacuum at 45 C to dryness, dissolved in 2ml ethanol and analysed by TLC and gas chromatography (GC). TLC was performed on silica gel plates (same as before) with chloroform/acetic acid/water (30:35:5) and n-butanol/acetone/water (4:5:1). Visualisation of spots was achieved with (i) 0.2% (w/v) naphtoresorcinol in ethanol:20% H2SO4 (1:1 v/v) and (ii) 1% a-naphtol:20% H2SO4 (1:1 v/v). GC of carbohydrates was carried out by dissolving hydrolysates, together with a 0.1ml solution of D-mannitol (Merck) as internal standard, in 80% ethanol (1.0mg/ml^-1). The mixture was evaporated to dryness under vacuum and treated with 0.5ml N,O-bistrimethylsilylacetamide in sealed vials. After heating at 100 C for 1 hr, 1.0ml of the mixture was injected into a Carlo Erba Science apparatus equipped with a 25m x 0.25mm WCOT column packed with DB-5 silicone oil. GC conditions were the same as for fatty acid analysis except for the initial temperature (110 C). Amino acids of NaOH soluble proteins were determined after extraction of mycelia with 0.5M NaOH followed by treatment with 15% (w/v) trichloracetic acid. Identification and quantitative analysis of amino acids were carried out after acid hydrolysis of the proteins with 6M HCl. Ion exchange chromatography was performed at 570 and 440 nm. Acid hydrolysis was performed by mixing protein samples (10mg) with 6M HCl in sealed vials and heating at 100 C for 20hrs. Reaction mixtures were filtered, evaporated under vacuum at 45 C, dry residues dissolved in 1.0ml methanol and analysed by TLC on silica gel with ethyl-methyl ketone/ ethanol/ NH3 (1:1:1). The plates were sprayed with a 1% (w/v) solution of ninhydrine in ethanol. Quantitative analysis of amino acids was carried out with a Biotronic LC-600 apparatus at 570 and 440nm under standard instrumental conditions. ------------------------------------------------------------- STRAIN SUBSTRATE MYCELIUM AERIAL MYCELIUM SPORULATION ------------------------------------------------------------- 1000 (H I) pink light pink good 2494 (H II-3-9) light pink light pink good H-1 cherry light pink poor H-2 yellow light pink poor H-3 lemon light pink poor H-4 cherry light pink poor H-13 cherry light pink poor H-14 red light pink poor H-72 yellow white good H-33 pink light pink good H-II-29 light yellow white poor H-f pink light pink good ------------------------------------------------------------Table 1. Characteristics of Streptomyces spectabilis strains.
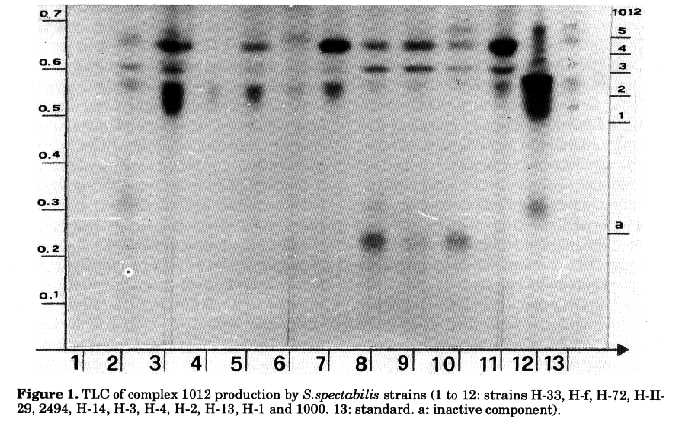
Cultural characteristics of the different strains of S.spectabilis are summarised in Table 1. Antibiotic complex 1012. The complex consists of three major components and two present in lower amounts (Fig. 1). The antibiotic complex inhibits Escherichia coli strain 26 and Bacillus subtilis strain 6633. Complex 1012 (Ivanova et al., 1991) does not react with Sakaguchi, Fehling and Benedict reagents and with anisic aldehyde. Positive reactions are observed with concentrated sulphuric acid and iodine vapour. Spectra of the components in methanol show (UV and the visible regions) two absorption maxima at 210 and 223nm. Antibiotic 1012 differs from the streptovaricin complex whose absorption maxima are at 245, 264, 320 and 430 nm. Spectomycin A2 shows maxima at 215, 260 and 399nm, spectomycin B1 at 224, 269 and 430nm, while spectinomycin and deoxymonomycins exhibit end absorption only.
--------------------------------------------------------------
FATTY STRAINS
ACIDS*
1000 2494 H-2 H-3 H-4 H-13
--------------------------------------------------------------
n-C9:1 - - - 0.22 - -
n-C9:0 0.29 - - 0.52 - 0.36
i-C10:0 - - - - - 0.15
n-C10:0 - - - - - 0.18
n-C11:1 - - - - - 0.14
n-C11:0 - 0.18 - 0.25 - 0.23
i-C12:0 0.48 0.20 0.48 0.30 0.71 0.99
n-C12:1 - - 0.14 0.48 0.45 -
n-C12:0 - - 0.41 0.45 1.31 -
a-C13:0 - 0.17 - 0.43 0.46 0.18
i-C13:0 - 0.19 - 0.99 0.82 1.88
n-C13:1 0.70 0.74 0.58 - - 0.52
n-C13:0 1.00 1.02 0.77 - - 0.88
i-C14:0 14.29 6.60 13.55 6.90 15.8717.34
n-C14:1 1.76 - - - - -
n-C14:0 1.19 1.25 1.52 3.36 1.76 2.35
a-C15:0 6.39 20.91 15.53 12.81 10.96 11.25
i-C15:0 20.32 24.51 19.91 31.67 19.50 16.97
n-C15:1 1.62 0.18 0.89 - - 0.22
n-C15:0 0.98 1.32 - 1.14 0.67 0.99
i-C16:0 4.49 2.97 4.80 2.56 4.02 3.38
n-C16:1 23.17 16.35 24.15 18.43 24.39 24.14
n-C16:0 4.36 4.01 4.96 2.29 6.59 5.70
x 4.92 4.97 4.09 5.07 7.50 4.74
a-C17:0 1.79 2.10 1.18 0.22 0.54 0.85
i-C17:0 1.62 1.98 1.53 0.95 1.56 1.14
n-C17:1 3.00 1.67 0.95 - 0.73 0.69
n-C17:0 5.15 0.94 3.32 - - 2.33
i-C18:0 - - - 0.53 2.90 0.60
x - - 0.09 - 1.09 -
n-C18:1 - 4.67 0.16 0.41 - 0.42
n-C18:3 - - - 0.37 - -
n-C18:0 - 2.61 0.17 0.19 - 0.14
--------------------------------------------------------------
FATTY STRAINS
ACIDS*
H-14 H-72 H-33 H-II-29 H-f
----------------------------------------------------------
----
n-C9:1 - - - - -
n-C9:0 0.10 - - - -
i-C10:0 0.09 - - - -
n-C10:0 - - - - -
n-C11:1 0.08 - 0.20 - 0.34
n-C11:0 0.09 - - - -
i-C12:0 0.64 0.61 - - -
n-C12:1 - 0.17 0.16 - -
n-C12:0 - 0.47 0.15 - -
a-C13:0 0.15 0.53 - - -
i-C13:0 0.42 1.20 - - -
n-C13:1 0.44 - - - 0.30
n-C13:0 0.82 - - - -
i-C14:0 14.52 10.84 10.83 4.57 3.30
n-C14:1 - 0.06 - 2.47 -
n-C14:0 1.17 1.43 0.58 2.26 0.66
a-C15:0 12.70 11.91 10.27 15.38
31.44
i-C15:0 21.20 23.30 17.05 -
14.98
n-C15:1 0.13 - 1.10 1.61 1.14
n-C15:0 1.34 0.47 - 0.81 -
i-C16:0 4.20 6.53 9.05 2.86 2.59
n-C16:1 26.74 20.12 30.61 21.40
13.99
n-C16:0 1.79 9.17 2.03 2.18 1.48
x 4.61 3.16 3.57 12.02 3.85
a-C17:0 0.88 1.54 0.25 1.95 0.60
i-C17:0 1.55 2.24 1.38 2.16 5.55
n-C17:1 1.12 - 4.68 4.17 3.42
n-C17:0 4.10 - 0.84 2.13 4.92
i-C18:0 0.71 3.59 3.65 7.90 6.20
x 0.09 0.17 1.35 2.26 1.50
n-C18:1 0.14 0.63 0.58 2.26 1.06
n-C18:3 - 0.41 - - -
n-C18:0 0.09 0.18 0.62 4.31 1.40
--------------------------------------------------------------
Table 2. Fatty acid composition of S.spectabilis
lipids. Data are expressed as percentages of the total peak
area on GC of methyl esters. ^*The first number indicates
carbon atoms and the second corresponds to the double bonds; n
= normal acid; - = iso acid; a = anteiso acid; x
= unidentified fatty acids.
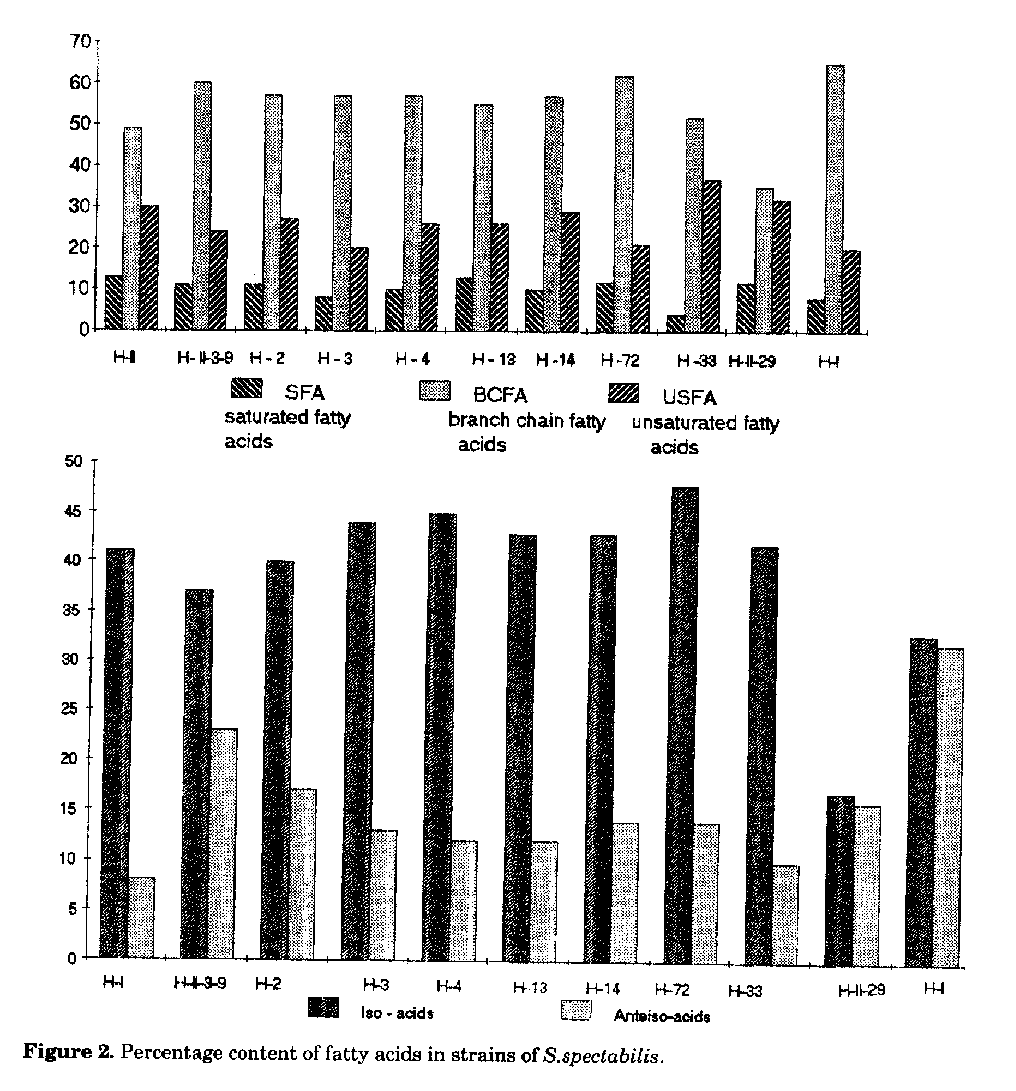
The parent strain 2494 (H-II-3-9) synthesises the two main components 1012-2, 1012-4, while the parent strain 1000 (H-I) produces the entire antibiotic complex, though in very low concentrations. Strains H-1 and H-3 synthesise as main components antibiotics 1012-2, 1012-3 and 1012-4, strains H-2 and H-4 components 1012-3 and 1012-4. Components 1012-2, 1012-3, 1012-4 and 1012-5 are produced in a nearly equal ratio by mutants H-13 and H-14. Strain H-33 shows no activity, while the strain H-II-29 forms only antibiotic 1012-2 in low concentrations. Strain H-72 produces the whole antibiotic complex 1012 and displays the highest activity (400-600g/ml). Strain H-f, resulting from protoplast fusion, is also an active producer, however with a lower activity than that of H-72. It synthesises components 1012-2, 1012-3, 1012-4 in nearly equal ratios. Lipids. The lipids of the strains investigated contain several fatty acids with 9 to 18 C atoms. The main components determined are iso and anteiso fatty acids with 14-18 C atoms; unsaturated acids with C 16:1 (hexadecenoic acid), C 17:1 (heptadecenoic acid) and saturated fatty acids with n-C 16:0 (palmitic acid). Fatty acid composition and percent ratio of saturated, iso, anteiso and unsaturated fatty acids of all the strains are shown in Table 2 and in Fig.2. Compared to S.spectabilis strain 1000, strain 2494 is characterised by a larger amount of unsaturated fatty acids; branched-chain fatty acids also predominate in the latter strain. The anteiso acid content in strain 1000 is about 15% lower than that of 2494. The quantitative ratio between saturated, iso, anteiso and unsaturated fatty acids in mutants obtained following streptovaricin treatment is almost the same (1:6:3), indicating that streptovaricin treatment does not affect fatty acids biosynthesis. More significant differences in the amounts of fatty acids are observed in strains obtained by UV-radiation and in fusant H-f. Strain H-72 synthesises a smaller amount of unsaturated fatty acids (20%), whereas H-33 shows a decrease (4.2%) in the amount of saturated fatty acids with respect to the unsaturated ones (37%). The iso and anteiso fatty acid ratio in strain H-II-29 and in H-f is 1:1, while in the remaining strains it is 4:1.
Carbohydrates of polysaccharides. Carbohydrates of the various strains have the same qualitative composition (Tab. 3 and 4).
-------------------------------------------------------------
Carbohydrate R[f]
A B
-------------------------------------------------------------
Unknown 0.19 0.26
Unknown 0.21 0.29
Galactose 0.26 0.33
Glucose 0.28 0.40
Mannose 0.31 0.44
Arabinose 0.34 0.47
Xylose 0.38 0.55
Table 3. R[f] values (TLC) of carbohydrates following acid hydrolysis of polysaccharides. Developing systems: A = CHCl3-AcOH-H2O (30:35:5), B = n-BuOH-Acetone-H2O (4:5:1). TLC and GC studies indicate the presence of xylose, arabinose, mannose, galactose, glucose and of two not yet identified carbohydrates. Rhamnose and amino sugars have not been found. More significant differences in the concentration of some carbohydrates can be detected in some strains. H-72 has a xylose:arabinose:mannose: galactose:glucose ratio of 1:1:1:1:0.5. The highest xylose and arabinose contents are found in strains H-33 and H-I.
-------------------------------------------------------------- CARBO- STRAINS HYDRATE 1000 2494 H-1 H-2 H-3 H-4 -------------------------------------------------------------- Xylose 2.81 2.74 5.54 0.30 1.04 1.09 Arabinose 1.00 2.70 4.25 0.22 0.60 0.26 Mannose 1.32 3.24 0.87 0.78 0.40 0.19 Galactose 0.71 1.60 1.32 0.24 0.73 0.30 Glucose 0.56 1.90 1.33 1.13 0.47 0.22 -------------------------------------------------------------- CARBO- STRAINS HYDRATE H-13 H-14 H-72 H-33 H-II-29 H-f -------------------------------------------------------------- Xylose 1.31 0.48 1.23 4.92 2.54 2.27 Arabinose 1.26 0.25 1.16 4.50 1.75 3.33 Mannose 3.80 1.02 1.76 0.89 4.63 6.11 Galactose 1.67 0.35 0.91 1.04 2.13 5.54 Glucose 0.57 0.32 0.45 1.24 0.62 0.31 --------------------------------------------------------------Table 4. Concentration (mol/100mg dry substance) of carbohydrates, as TMSi derivatives, in the polysaccharides of S.spectabilis strains.
--------------------------------------------------------------
STRAIN
AMINO ACID 1000 2494 H-3 H-72
--------------------------------------------------------------
Aspartate 87.1 51.7 99.5 81.3
Threonine 60.6 40.2 76.6 61.0
Serine 55.5 34.1 64.2 47.0
Glutamate 224.2 155.7 175.7 204.8
Proline 51.0 - 64.7 -
Glycine 256.7 169.4 173.3 331.0
Alanine 253.0 182.0 239.3 342.7
Cysteine - - - 66.3
Valine 119.4 82.3 138.0 146.6
Methionine 102.4 94.0 71.0 155.4
Isoleucine 50.2 40.3 54.4 48.0
Leucine 100.7 68.2 105.1 79.5
Tyrosine 17.6 17.5 18.2 25.1
Phenylalanine 29.9 21.9 32.1 36.6
Histidine 77.6 109.0 110.4 123.3
Lysine 46.8 54.0 73.5 65.5
Arginine 62.8 - - -
Diaminopimelate 12.1 18.0 20.5 21.0
Table 5. Concentration (nmol/mg protein) of amino acids in the proteins of some S.spectabilis strains.
Amino acids of basic and acid soluble proteins. Eighteen amino acids were detected (Table 5). All strains show a high content of methionine, histidine, glutamate, glycine, alanine, valine and leucine, while that of tyrosine and DL-2,6-diaminopimelic acid is quite low. Proteins exhibit qualitative and quantitative differences in the composition and concentration of the amino acids. Mutant H-72 contains cysteine, absent in other strains, but no proline or arginine. Proline is also present in proteins of parent strain 2494, while arginine is only detected in H-I. In comparison to the other strains, mutant H-72 has the highest concentration of valine, glycine, alanine, methionine and histidine. DISCUSSION The present investigation was carried out in order to establish a possible correlation between the synthesis of antibiotic complex 1012 and cell components (lipids, polysaccharides and proteins). Mutant H-72 shows the highest antibiotic activity and synthesises the entire antibiotic complex 1012. In comparison with the other strains, H-72 shows larger amounts of fatty acids, amino acids and carbohydrates. As far as the fatty acid composition is concerned, the mutant synthesises a high percentage of unsaturated and of branched-chain fatty acids. In contrast to the rest of the strains, the biosynthesis of iso considerably prevails over the that of anteiso fatty acids (ratio 3.5:1). The qualitative composition of polysaccharides is similar, however the xylose, arabinose, mannose, galactose, glucose ratio is different from that of the other strains. H-72 is the only strain containing cysteine but no proline. It also shows the highest content of glutamate, glycine, alanine and histidine. Fusant H-f has a lower antibiotic activity and synthesises components 1012-2, 1012-3 and 1012-4. A characteristic of the strain is the high percentage of fatty acids with a branched chain, however it differs from H-72 in the quantitative iso/ anteiso fatty acid ratio. In addition strain H-f shows a high content of mannose and galactose. REFERENCES Christie, W.W. (1973). Lipid Analysis. Pergamon Press, Oxford Dietz, A., C.DeBoer, R.M.Smith, P.Siminoff, G.A.Boyack & G.B.Whitfield (1963). Antibiotic streptovaricin and process for its production. US Patent 3 116 202
Gauze, G.F., T.P.Preobrazhenskaya, L.P. Sveshnikova, L.P.Terechova & T.S.Maximova (1983). Key to the Actinomycetes. Nauka Publ., Moscow
Ivanova, V., N.Vesselinova & R.Gesheva (1987). Isolation and identification of antibiotic complex 1011 from Streptomyces sp. 1011. Acta Microbiol.Bulg., 21: 47-50
Ivanova, V., N.Vesselinova & R.Gesheva (1989). Prodigiosin-like pigments from Streptomyces sp. 1011. V Int.Conf.Chem. & Biotech.Biol. Active Natural Products, 1: 412-417
Ivanova, V., N.Vesselinova & R.Gesheva (1991). Method for production of antibacterial antibiotic. Bulgaria Patent N. 84704
Mason, D.J., A.Dietz & R.M.Smith (1961). Actinospectacin, a new antibiotic. Discovery and biological properties. Antib.Chemother., 2: 118-133
Staley, A.L. (1990). Isolation, structure, synthesis and biosynthesis of selected bioactive microbial metabolites. Abstr.Ph.D. Diss., Univ. of Illinois at Urbana-Champaign
Thompson, C.J., J.M.Ward & D.A.Hopwood (1982). Cloning of antibiotic resistance and nutritional gene in Streptomyces. J.Bacteriol., 115: 668-677
Yamazaki, H. (1968a). Studies on antimicrobial substance B 44P (streptovaricin), produced by a strain of Actinomycetales. I. Production, extraction and characteristics of substance B 44P and identity of substance with streptovaricin. J. Antib., 21: 204-208
Yamazaki, H. (1968b). Studies on antimicrobial substance B 44P (streptovaricin) produced by a strain of Actinomycetales. II. Microbial and pharmacological studies. J.Antib., 21: 209-221. Copyright 1995 CETA
The following images related to this document are available:Halftone images[ac95008a.gif]Photo images[ac95008a.jpg]Line drawing images[ac95008b.gif] |
| |||||||||
{kind=link}
{kind=link}