 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
DISTRIBUTION OF ANTIBIOTIC-PRODUCING MICROBISPORA STRAINS IN SOILS WITH DIFFERENT pHs M. HAYAKAWA, K. ISHIZAWA, T. YAMAZAKI and H. NONOMURA DepT of Applied Chemistry and Biotechnology, Yamanashi University, Takeda-4, Kofu 400, Japan
Code Number: AC95013
Sizes of Files:
Text: 16K
Graphics: Line Drawings (gif) 12K
ABSTRACT. A total of 439 cultures of Microbispora
spp., freshly isolated from 117 different soil samples, were
investigated for their antimicrobial activity using a humic
acid-containing medium. Eighty-seven (20%) isolates were
active against Staphylococcus aureus and only 12
(3%) against Aspergillus niger. The incidence of
antibiotic producers increased with soil pH.
Species of the genus Microbispora produce longitudinal pairs of spores on the aerial mycelium (Nonomura and Ohara, 1957). Although microbisporae represent only a minor component of the actinomycete population in soil (Hayakawa et al., 1988), it has been suggested that they may play a significant role in the breakdown of the recalcitrant organic polymers, such as cellulose and xylan (Waldron et al., 1986; Ball and Mc Carthy, 1988). Microbisporae have also been recognised as a source of antibiotics and other bioactive compounds, such as phenazines (Gerber and Lechevalier, 1964; Tanabe et al. 1995), the antifungal antibiotic Sch 31828 (Patel et al., 1988), cochinmicins (Lam et al., 1992), and angelmicins (Uehara et al., 1993). In the search for new antibiotics, the probability of isolating novel producers is enhanced by screening substrates harbouring a rich flora of antagonistically active organisms (Kutzner, 1989). A number of researchers have pointed out a possible correlation between occurrence of antagonistic actinomycetes and nature of the isolation soil. Rouatt et al. (1951) found a greater percentage of active streptomycetes in the rhizosphere than in the surrounding soil. According to Khan and Williams (1975) antifungal activity was predominant among acidophilic streptomycetes common in acid soils. On the other hand, acidoduric streptomycetes inhabiting forest soils showed greater activity than neutrophiles against Gram-negative bacteria (Nkanga and Hagedorn, 1978). Recently numerous cultures of non-streptomycetes, such as maduromycetes and actinoplanetes, have been isolated from soils and screened for new bioactive compounds (Okami and Hotta, 1988). Little attention however has been paid to the effect of soil types on the occurrence of antagonistic non- streptomycetes. The present paper describes the results of an investigation on the occurrence of antibiotic-producing microbisporae from Japanese soils characterised by different pHs.
MATERIALS and METHODS
Soil samples. A total of 117 soil samples, mainly from cultivated fields, were collected in different locations in Japan. Samples were sieved (2mm mesh) and air- dried at room temperature for 7dd. Bacterial counts were carried out by dilution plate methods (Nonomura and Takagi, 1977). Chemical characteristics of the samples were analysed according to standard procedures (Hayakawa and Nonomura, 1987). Isolation of microbisporae. Soil samples were subjected to dry heat treatment (1h at 120 C; Nonomura and Ohara, 1969). Logarithmic dilutions were prepared in sterile tap water and 0.2ml aliquots spread onto five plates containing a humic acid-vitamin medium (Hayakawa and Nonomura, 1987) supplemented with cycloheximide (50 mg/l; Williams and Davies, 1965). Plates were incubated at 30 C for 3 weeks and actinomycete colonies examined directly under a light microscope equipped with a 40x long working distance objective (Model ULWDCDPlan, Olympus, Tokyo). Microbispora colonies were recognised by the presence of a well-developed aerial mycelium with longitudinally paired spores, closely arranged or borne at relatively longer intervals (Hayakawa et al., 1991). For each soil sample, three to five randomly selected colonies were subcultured onto oatmeal- YGG agar (Hayakawa et al., 1982). A total of 439 Microbispora cultures was isolated from the soil samples. Taxonomic determinations. Isolates were grown on oatmeal-Y agar (Nonomura and Ohara, 1960) at 30 C for 3 weeks and checked by light microscopy. To complement generic identification, 30 representative isolates were further investigated for a range of other taxonomic characteristics. Fine spore morphology was determined by scanning electron microscopy (SEM) (JSM T100, Jeol Ltd., Tokyo), following a previously described specimen preparation method (Hayakawa et al., 1991). The biomass was analysed for sugar and diaminopimelic acid (DAP) isomer composition (Hasegawa et al., 1983). Antimicrobial activity. The ability of the isolates to produce antibiotic substances was ascertained by the conventional paper disc-plate method (Seto, 1992). Isolates were cultivated in submerged culture in test tubes in AP broth (pH 7.0) supplemented with humic acid (1g/l) dissolved in 0.2N NaOH (Hayakawa and Nonomura, 1987). AP broth contains (g/l) glucose (10), soluble starch (10) Difco yeast extract (5), peptone (3), soybean flour (10; Sigma Chemical Co., St.Louis, MO, USA), NaCl (2), CaCO3 (3) and distilled water. Test tubes were incubated with reciprocal shaking (240 strokes/min) at 30 C for 12dd. Fermentation broths were tested for against Staphylococcus aureus IFO 3061 and Aspergillus niger ATCC 9642.
RESULTS and DISCUSSION
Generic identification. Morphological and chemical properties of the 30 isolates were consistent with their placement in the genus Microbispora (Nonomura and Ohara, 1957).
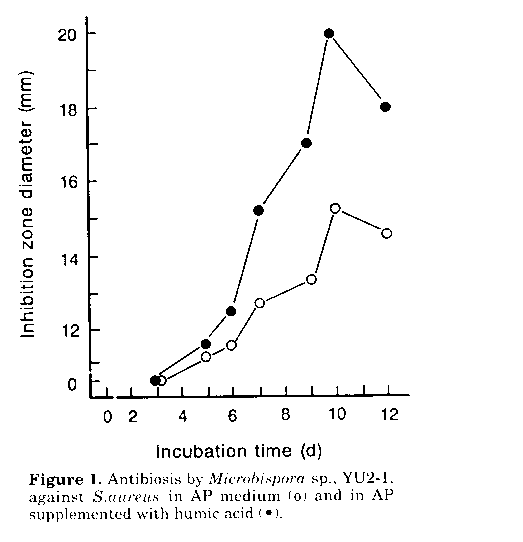
Figure 1. Antibiosis by Microbispora sp., YU2- 1, against S.aureus in AP medium and in AP supplemented with humic acid.
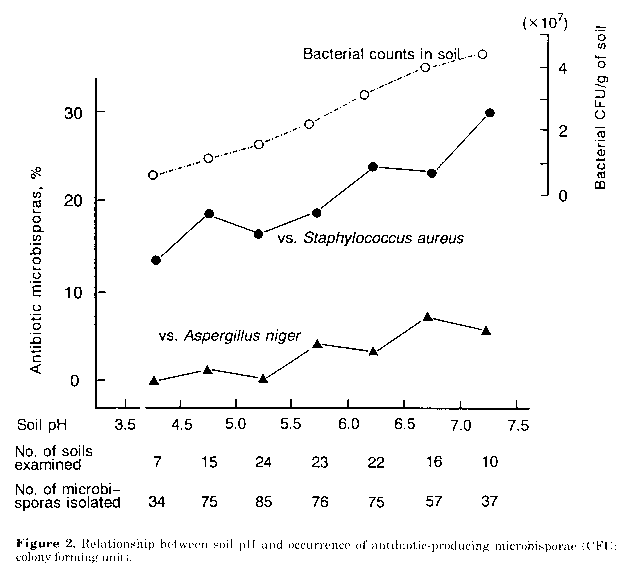
Figure 2. Relationship between soil pH and occurrence of antibiotic-producing microbisporae (CFU: colony forming unit).
Stimulating effects of humic acid on antibiotic productivity. Investigations were carried out, in the light of previous reports (Martin et al., 1976) on the role of humic acid on growth and metabolism of actinomycetes. As shown in Fig. 1 production of antibiotic substance(s) by Microbispora sp., YU2-1, was markedly enhanced by humic acid (1.0g/l) in the fermentation medium. A similar effect was observed with four other strains of Microbispora and M.aerata P-132, a producer of iodinin (Gerber and Lechevalier, 1964). In this study the mechanism of the stimulatory effect of the humate, as well as the nature of the antimicrobial substances produced by the Microbispora isolates, were not investigated. However results suggest that humate- containing fermentation media can be used to stress the potential antibiotic activity of microbisporae. Occurrence of antibiotic-producing microbisporae in soils. When tested on humic acid supplemented fermentation medium, 92 (21%) of the 439 Microbispora isolates showed antimicrobial activity. Most strains (20%) inhibited S.aureus, while only 12 (3%) were active against A.niger. The incidence of the microbisporae active against S.aureus showed a tendency to increase with the pH value of the soil source (Fig. 2). Correspondingly a comparatively higher percentage of organisms active against A.niger was found among isolates from soils with a pH range of 5.5-7.5. Results obtained indicate that neutral to slightly alkaline soils, also harbouring a quantitatively rich flora of eubacteria (Fig. 2), may be the best source of antibiotic- producing microbisporae. However additional work is needed to explain the occurrence of the antibiotic microbisporae in acid soils, since the pH value of the isolation medium used was 7.2. Using natural and man-made acid soils and adjusting the pH of culture media to the same value as that of the soil, Williams et al. (1971) and Khan and Williams (1975) were able to isolate acidophilic streptomycetes with antifungal activity. In neutral to alkaline habitats, it seems likely that eubacteria are microbisporae's main competitors. Therefore it is possible to assume that their ability to produce antibacterial antibiotics may endow them with advantages in the colonisation of, and survival in, such environments. However it should be noted that there is no direct evidence of the presence of antibiotics in natural soils (Williams et al., 1989).
ACKNOWLEDGEMENTS. We are grateful to Dr. T. Beppu for his encouragement. We thank Mrs.J.Rogers for revising the manuscript. This research was supported by a grant from the Waksman Foundation of Japan, Inc.
REFERENCES
Ball, A.S. & A.J.McCarthy (1988). Saccharification of straw by actinomycete enzymes. J.gen. Microbiol., 134: 2139-2134
Gerber, N.N. & M.P.Lechevalier (1964). Phenazines and phenoxazinones from Waksmania aerata sp. nov. and Pseudomonas iodina. Biochem., 3: 598-602
Hasegawa, T., M.Takizawa & S.Tanida (1983). A rapid analysis for chemical grouping of aerobic actinomycetes. J.Gen.Appl.Microbiol., 29: 319-322
Hayakawa, M., S.Iino & H.Nonomura (1982). Heavy metal resistance and melanoid pigment production in the streptomycete flora of copper-polluted vineyard soils. Hakkokogaku, 60: 1-9 Hayakawa, M. & H.Nonomura (1987). Humic acid-vitamin agar, a new medium for the selective isolation of soil actinomycetes. J.Ferment. Technol., 65: 501-509
Hayakawa, M., K.Ishizawa & H.Nonomura (1988). Distribution of rare actinomycetes in Japanese soils. J.Ferment.Technol., 66: 367-373
Hayakawa, M., T.Sadakata, T.Kajiura & H. Nonomura (1991). New methods for the highly selective isolation of Micromonospora and Microbispora from soil. J.Ferment.Bioeng., 72: 320-326
Khan, M.R. & S.T.Williams (1975). Studies on the ecology of actinomycetes in soil. VIII. Distribution and characteristics of acidophilic actinomycetes. Soil Biol.Biochem., 7: 345-348
Kutzner, H.J. (1981). The family Streptomycetaceae. In: M.P.Starr, H.Stolp, H.G.Trper, A. Balows & H.G.Schlegel (eds.) The Prokaryotes. A Handbook on Habitats, Isolation and Identification of Bacteria. Springer-Verlag, New York, pp. 2028-2090
Lam, Y.K.T., D.L.Williams, Jr., J.M.Sigmund, M.Sanchez, O.Genilloud, Y.L.Kong, S.Stevens-Miles, L.Huang & G.M.Garrity (1992). Cochinmicins, novel and potent cyclodepsipeptide endothelin antagonists from a Microbispora sp. I. Production, isolation, and characterization. J.Antibiot., 45: 1709-1716
Lechevalier, M.P. & H.Lechevalier (1970). Chemical composition as a criterion in the classification of aerobic actinomycetes. Int.J.Syst.Bacteriol., 20: 435- 443
Martin, J.P., Z.Filip & K.Haider (1976). Effect of montmorillonite and humate on growth and metabolic activity of some actinomycetes. Soil Biol.Biochem., 8: 409- 413
Nkanga, E.J. & C.Hagedorn (1978). Detection of antibiotic- producing Streptomyces inhabiting forest soils. Antimicr.Ag.Chemother., 14: 51-59
Nonomura, H. & Y.Ohara (1957). Distribution of actinomycetes in soil. II. Microbispora, a new genus of Streptomycetaceae. J.Ferment.Technol., 35: 307-31 1
Nonomura, H. & Y.Ohara (1960). Distribution of actinomycetes in soil. IV. The isolation and classification of the genus Microbispora. J. Ferment.Technol., 38: 401-405 Nonomura, H. & Y.Ohara (1969). Distribution of actinomycetes in soil. VI. A culture method effective for both preferential isolation of Microbispora and Streptosporangium in soil (Part I). J. Ferment.Technol., 47: 463-469
Nonomura, H. & S.Takagi (1977). Distribution of actinoplanetes in soils of Japan. J.Ferment. Technol., 55: 423-428
Okami, Y. & K.Hotta (1988). Search and discovery of new antibiotics. In: M.Goodfellow, S.T.Williams & M.Mordarski (eds.) Actinomycetes in Biotechnology. Academic Press, New York, pp. 33-67
Patel, M., M.Conover, A.Horan, D.Loebenberg, J.Marquez, R.Mierzwa, M.S.Puar, R. Yarborough & J.A.Waitz (1988). Sch 31828, a novel antibiotic from a Microbispora sp.: taxonomy, fermentation, isolation and biological properties. J.Antibiot., 41: 794- 797
Seto, H. (1992). Antibiotics. In: T.Beppu, A.Nomoto & T.Ota (eds.) Microbiological Methods (in Japanese). Tokyo Kagaku Doujin, pp. 150-152
Tanabe, I., M.Kuriyama & H.Nonomura (1995). Production of C2-symmetrical phenazines by some actinomycetes. J.Ferment.Bioeng., 79: 384-386
Uehara, Y., P-M.Li, H.Fukazawa, S.Mizuno, Y.Nihei, M.Nishio, M.Handa, C.Yamamoto, T.Furumai & T.Oki (1993). Angelmicins, new inhibitors of oncogenic src signal transduction. J. Antibiot., 46: 1306- 1308
Waldron, C.R., C.A.Becker-Vallone & D.E. Eveleigh (1986). Isolation and characterization of a cellulolytic actinomycete Microbispora bispora. Appl.Microbiol.Biotechnol., 24: 477-486
Williams, S.T. & F.L.Davies (1965). Use of antibiotics for selective isolation and enumeration of actinomycetes in soil. J.gen.Microbiol., 38: 251-262
Williams, S. T., F.L.Davies, C.I.Mayfield & M.R.Khan (1971). Studies on the ecology of actinomycetes in soil. II. The pH requirements of streptomycetes from two acid soils. Soil Biol. Biochem., 3: 187-195
Williams, S.T., M.Goodfellow & G.Alderson (1989). Genus Streptomyces Waksman and Henrici 1943. In: S.T.Williams, M.E.Sharpe & J.G. Holt (eds.) Bergey's Manual of Systematic Bacteriology. The Williams & Wilkins Co., Baltimore, Vol. 4, pp. 2452-2492. Copyright 1995 C.E.T.A
The following images related to this document are available:Line drawing images[ac95013b.gif] [ac95013a.gif] |
| |||||||||
{kind=link}
{kind=link}