 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ultrastructural Investigations On Micromonospora And Rhodotorula Species Grown In Pure And Mixed Culture
V. GESHEVA, R. GESHEVA, N. KOSTRIKINA* and V. BIRJUZOVA* Institute of Microbiology, Bulgarian Academy of Sciences, 1113 Sofia Bulgaria and *Institute of Microbiology, Russian Academy of Sciences, Moscow, Russia
Code Number: AC96002
Size of Files:
Text: 15.5K
Graphics: Photographs (jpg) - 500KABSTRACT. The ultrastructure of Rhodotorula and Micromonospora species grown in mixed culture was investigated. Cells of Rhodotorula showed increased cell wall thickness and shortening of fibrils. The results may be imputed to trophic effects.
Under natural conditions microorganisms grow and develop in communities. Different forms of interrelation (neutralism, symbiosis, antagonism, parasitism, predatoriness) may conse- quently ensue. Interactions are variable and affected by environmental conditions. In the study of fungal and streptomycete relationships, great attention has been paid to the antagonism by actinomycetes (Skinner, 1956; Rehm, 1958; Grigorov and Gesheva, 1960; Shen et al., 1977; Rotzrock and Gottlieb, 1981; Smith et al., 1990; Inamori et al., 1990; Takanori and Takanashi, 1993; Mostafa-Mahmoud, 1993). In the search for an- tagonists against phytopathogens, the role of volatile metabolites produced by streptomycetes in soil fungistasis has also been investigated (Hora and Baker, 1972; Fries, 1973; Gupta and Tandom, 1977; Rai et al., 1981; Herrington et al., 1987). So far however interaction mechanisms between fungi and actinomycetes have not been elucidated. Recently lytic activity on fungal hyphae and effects on reproduction structures have been reported (Rai and Upadhyay, 1978; Sabaou et al., 1981, 1983; Sabaou and Bounaga, 1987; Sugahara et al., 1992; Mostafa-Mahmoud, 1993). In the present paper ultrastructural investigations on mixed cultures of Micromonospora and Rhodotorula are illustrated.
MATERIALS and METHODS
Organisms. Micromonospora sp., strain 220 of our collection, isolated from soil in Bulgaria, was grown on glucose- nitrate agar, mineral agar No. 1, maize agar No. 6 and peptone extract maize agar (Gauze et al., 1983). The strain pro- duces gentamicin and an unidentified antibiotic (unpublished results). The Rhodotorula sp. strain, a fresh soil iso- late, was grown on malt extract agar. Mixed cultures. Strain 220 was grown on mineral agar No. 1 for 10-14dd at 28 C and the yeast on malt extract agar for 2- 3dd at 28 C. Both organisms were streaked on mineral agar No. 1 and incubated for 10-14dd at 28 C. The association was maintained for 3 years and then examined by electron microscopy. Electron microscopy. Ten day old samples, grown in pure and in mixed culture (medium No. 6), were fixed in glutaraldehyde (5% v/v in phosphate buffer, pH 7.4) for 2.5hrs. Specimens were then transferred to a sucrose solution (0.2M) at 4 C for 12hrs. Postfixation was carried out in osmium tetroxide (3% w/v, same buffer). Rhodotorula cells were fixed according to Mller (Birjuzova et al., 1963) and also treated with uranylacetate (3% w/v in 30% ethyl alcohol) for 1h. Specimens were dehydrated in a graded alcohol series and acetone and embedded in Epon. Ultrathin sections were prepared with a LKB 4800 microtome and stained according to Reynolds (1963). Samples were examined with a JEM-100C electron microscope.
RESULTS and DISCUSSION
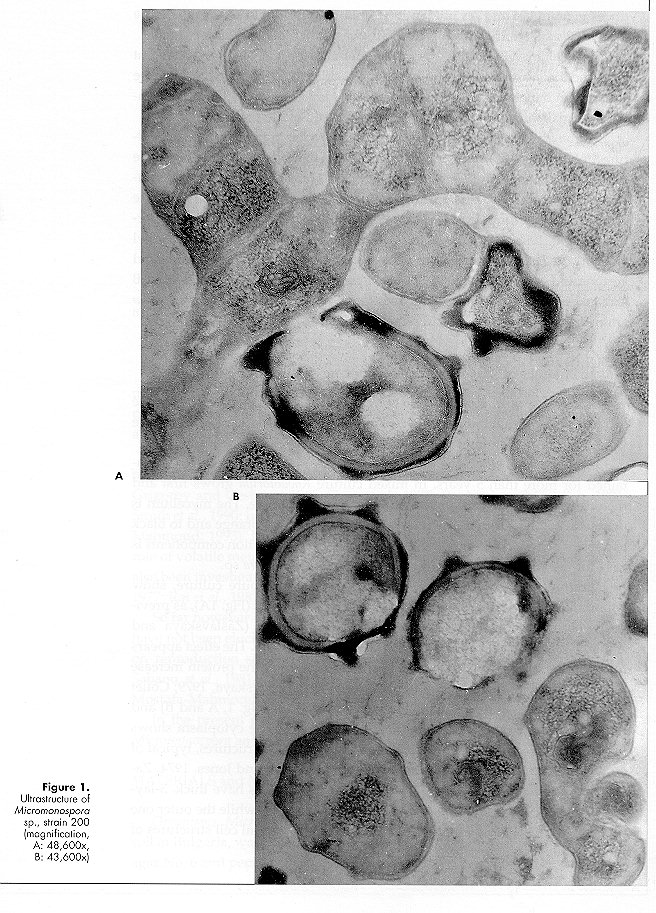
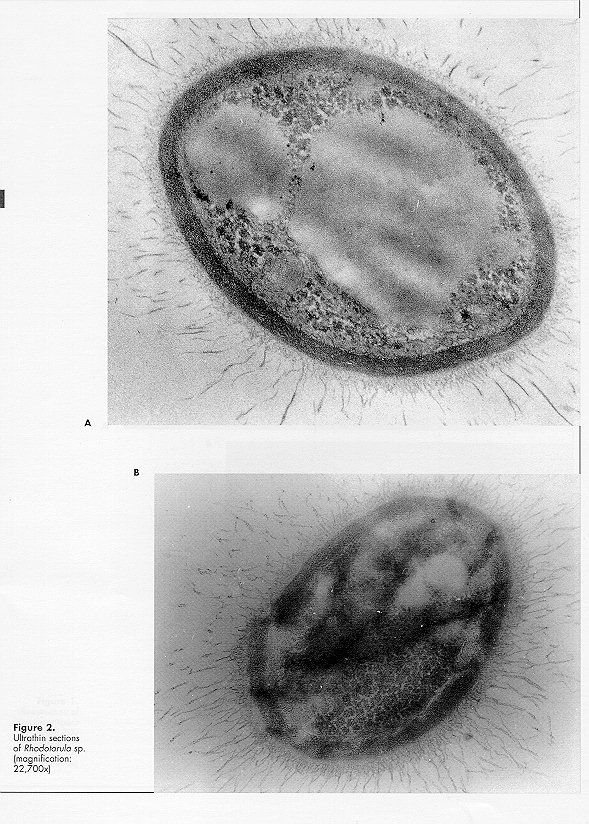
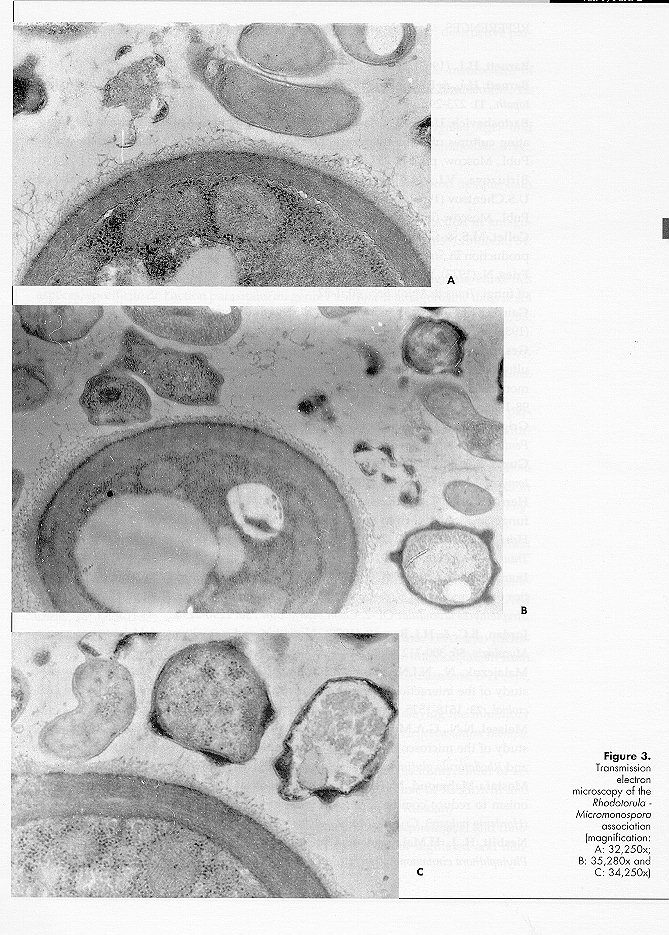
Micromonospora sp. strain 220 did not grow, or only poorly, on peptone maize extract and glucose nitrate agar. Mixed cultivation with the Rhodotorula isolate improved rate and amount of growth and sporulation of the actinomycete. On maize agar No. 6 the association is very stable and has been maintained for more than 5 years. In mixed culture Rhodotorula grows first and then Micromonospora appears on the yeast growth surface. The mycelium is orange in colour, turning progressively with age to dark orange and to black as a result of abundant sporulation. Isolation of the as- sociation components is easy for the yeast and more difficult for the Micromonospora sp. Ultrathin sections of Micromonospora sp., grown in pure culture, show abundant ribosomes, sometimes aggregated in polysomes (Fig. 1A), as previously observed for other antibiotic producing strains (Zaslavskaya and Zhukov, 1976; Zaslavskaya et al., 1977; Gesheva et al., 1984). The effect appears to be caused by the toxic activity of the antibiotic or by the protein increase following antibiotic production (Bartoshevich and Zaslavskaya, 1979; Collet and Jones, 1974). Cytoplasmic membranes are present (Fig. 1, A and B) and hyphal walls are typical of gram-positive organisms. The cytoplasm shows lipid inclusions and electron-transparent homogeneous structures, typical of strains producing biologically active metabolites (Collet and Jones, 1974; Zaslavskaya and Zhukov, 1976; Gesheva et al., 1984). Spores have thick, 3-layered walls; the inner layer, osmiophilic, forms the spines, while the outer one (lipidic monolayer) is membranous and smooth. All typical cell structures of the sporoplasm are present. Micromonospora sp. strain 220, grown in mixed culture, shows a similar morphology. The morphology (Fig. 2, A and B) of Rhodotorula sp., grown in pure culture, is typical (Meissell et al., 1962), with fib- rillar material present on the cell surface (Fig. 2 B). Malajczuk et al. (1977), investigating the association of Phytophthora cinnamomi with other organisms, showed that the fungus produces a mucopolysaccharide slime favouring the attachment of bacteria and actinomycetes. In the authors opinion the material is excreted as a reaction to the antibiotics produced by the latter organisms since it is absent when the fungus is grown in pure culture. During our investigations long fibrils on the cell surface of the yeast grown in pure culture were detected, while capsular material was much reduced in mixed culture. Polysaccharide fibrils were short and tended to disappear in the regions of contact with actinomycete hyphae (Fig. 3A), the capsular material being utilised by the Micromonospora strain. It appears therefore that the association between the two organisms is a trophic one, possibly a case of biotrophic or balanced parasitism (Barnett, 1963; Barnett and Binder, 1973; Jordan and Barnett, 1978). Yeast cells in mixed culture showed wall thickening (Fig. 3, B and C), as previously described (Malajczuk et al., 1977; Nesbitt et al., 1981), possibly as a defence mechanism against the presence of toxic metabolites. Other cytological changes were not observed.
REFERENCES
Barnett, H.L. (1963). Nature of mycoparasitism by fungi. Ann.Rev.Microbiol., 17: 1-14 Barnett, H.L. & F.L.Binder (1973). The fungal host parasite relationship. Ann.Rev.Phytopath., 11: 273-292
Bartoshevich, U.K. & P.L.Zaslavskaya (1979). Biosynthesis of antibiotics in differentiating cultures of actinomycetes and fungi. In: Ontogenesis of Microorganisms. Nauka Publ., Moscow, pp. 242-257 (in Russian)
Birjuzova, V.I., N.B.Borovechin, N.A.Gilev, A.C.Kiselov A.S.Tichonenko & U.S.Chentsov (1963). Electron-microscopic methods for studying biological samples. Nauka Publ., Moscow (in Russian)
Collet, M.S. & G.H.Jones (1974). Morphological changes accompanying actinomycin production in Streptomyces antibioticus. Ultrastr.Res., 46: 452-465
Fries, N. (1973). Effect of volatile organic compounds on the growth and development of fungi. Trans.Br.Mycol.Soc., 60: 1-22
Gauze, G.F., T.P.Preobrazhenskaya, M. Sveshnikova, L.P.Terechova, T.S.Maximova (1983) . Key to Actinomycetes. Nauka Publ., Moscow (In Russian)
Gesheva, V.I., N.A.Kostrikina, V.I.Birjuzova, N.S.Landau & N.S.Egorov (1984). The ultrastructural organization of Actinomyces rimosus and Actinomyces violocinereus in monocultures and in association producing exocellular proteases. Mikrobiologiya, 53: 98-102 (in Russian)
Grigorov, I. & R.Gesheva (1960). Interaction between Actinomyces globisporus and Penicillium citroroseum. Izv.Microbiol.Inst., 12: 59-65 (in Bulgarian)
Gupta, R.S. & R.H.Tandom (1977). Growth inhibition of fungi by volatiles from Streptomyces. Trans.Br.Mycol.Soc., 68: 438-439
Herrington, P.K., Y.E.Kreig & Y.E.Sheridan (1987). Methylvinyl ketone a volatile fungistatic inhibitor from Streptomyces griseoruber. Soil Biol.Biochem., 19: 509-512
Hora, T.S. & RBaker (1972). Soil fungistasis: microflora producing a volatile inhibitor. Trans.Br.Mycol.Soc., 59: 491-500
Inamori, J., H.Amino, M.Tsuboi, S.Jamaguchi & H.Tsuijbo (1990). Biological activities of racemomycin B, á-lysine rich streptothricin antibiotic, the main component of Streptomyces lavendulae OP-2. Chem.Pharm.Bull., 38: 2296-2298
Jordan, E.C. & H.L.Barnett (1978). Nutrition and parasitism of Melanospora zamiae. Mycologia, 50: 300-312
Malajczuk, N., N.J.Nesbitt & A.R.Glenn (1977). A light and electron microscope study of the interaction of soil bacteria with Phytophthora cinnamomi Rands. Can.J.Microbiol., 23: 1518-1525
Meissel, N.N., G.A.Medvedeva, V.I.Birjuzova & T.M.Volkova (1962). A comparative study of the microscopic and ultramicroscopic structure of cells of Saccharomyces vini and Rhodotorula glutinis. Mikrobiologiya, 31: 1011-1017 (in Russian)
Mostafa-Mahmoud, M. (1993). Biological control of Drechslera teres: Ability of antagonism to reduce conidia formation, coleoptile infection and leaf infection in barley (Hordeum vulgare). Cryp- tog.Mycol., 14: 287-295.
Nesbitt, H. J., H.Malajczuk & A.R.Glenn (1981). Bacterial colonization and lysis of Phytophthora cinnamomi. Trans.Br.Mycol.Soc., 77: 47-54
Rai, S. & R.R.Upadhyay (1978). Micromonospora globosa Krassilnikov: destructive parasite of Fusarium udum Butler. Microbios Lett., 31: 123-128
Rai, S., A.K.Srivasta & D.B.Singh (1981). Volatile and nonvolatile metabolites of actinomycetes and the growth of some litter decomposing fungi. Soil Biol.Biochem., 13: 75-76
Rehm, H.J. (1958). Untersuchungen ber das Verhalten von Aspergillus niger und einem Streptomyces albus Stamm in Mischkultur. Zbl.Bakt. Paras.Infektionskr. Hyg., II.Abt., 111: 111-115
Reynolds, R.S. (1963). The use of lead citrate at high pH as an electron opaque stain in electron microscopy. J.Cell Biol., 17: 208- 212
Rotzrock, C.S. & D.Gottlieb (1981). Importance of antibiotic production in antagonism of selected Streptomyces species in two soil-born plant pathogens. J.Antibiot., 34: 830-835
Sabaou, N. & N.Bounaga (1987). Actinomyc‚tes parasites de champignons: ‚tude des esp‚ces, specificit‚ de l'action parasitaire au genre Fusarium et antagonisme dans le sol envers Fusarium oxysporum f.sp. albedinis (Killian et Maire) Gordon. Can.J.Microbiol., 33: 445-451
Sabaou, N., M.Bennaceur & D.Bounaga (1981). Le palmier dattier et la fusariose. VIII. Action parasitaire d'un actinomyc‚te envers Fusarium oxysporum f.sp. albedinis. Ann.Microbiol., 132A: 319- 329
Sabaou, N., N.Bounaga & D.Bounaga (1983). Actions antibiotique mycolitique et parasitaire de deux actinomyc‚tes envers Fusarium oxysporum f.sp. albedinis et autres formae speciales. Can.J.Microoiol., 29: 194-199
Shen, B., S.J.Humble & J.L.Lockwood (1977). Parasitism of oospores of Phytophthora megasperma var. sojae, Phytophthora cactorum, Pythium sp. and Aphanomyces euteiches in soil by oomycetes, chytridiomycetes, hyphomycetes, actinomycetes and bacteria. Phytopathology, 67: 622-628
Skinner, F.A. (1956). Inhibition of the growth of fungi by Streptomyces spp. in relation to nutrient conditions. J.gen.Microbiol., 16: 2-6
Smith, S., A.Putnam & M.Nair (1990). In vitro control of Fusarium diseases of Asparagus officinalis L. with a Streptomyces or its polyene antibiotic faerifungin. J.Agr.Food Chem., 38: 1729-1733 Sugahara, I, T.Kimura, H.Inoue, T.Ohishi, K.Mase, J.Kawamura, H.Anano, J.Kitagawa & E.Eutani (1992). Isolation and properties of a fungilytic microorganism from mangrove sediments. Abstr. 6th Int.Symp.Microbiol. Ecol. (ISME 6), Barcelona, 6-11 Sept, 1992, p. 235
Takanori, S. & K.Takahashi (1993). A simplified method for assaying the inhibitory activity of actinomycete isolates against Rosellinia necatrix and Helicobasidium mompa. Bull.Nat.Inst. Sericult. Entomol.Sci., 7: 65-75
Zaslavskaya, P.L. & V.G.Zhukov (1976). Morphological changes in mycelium of active and nonactive variants off Actinomyces parvulus in the course of its growth and biosynthesis of actinomycin D. Mikrobiologiya, 45: 1035-1039 (in Russian)
Zaslavskaya, P.L., V.G.Makarevich & M.G. Slugina (1977). Morphological and functional investigations of Actinomyces aureofaciens in condi- tions of regulated and nonregulated fermentation. Mikrobiologiya, 46: 283-287 (in Russian) Figure 1. Ultrastructure of Micromonospora sp., strain 200 (magnification, A: 48,600x, B: 43,600x) Figure 2. Ultrathin sections of Rhodotorula sp. (magnification: 22,700x) Figure 3. Transmission electron microscopy of the Rhodotorula - Micromonospora association (magnification: 48,600x) Copyright 1996 C.E.T.A., The International Centre for Theoretical and Applied Ecology, Gorizia
The following images related to this document are available:Photo images[ac96002c.jpg] [ac96002b.jpg] [ac96002a.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}