 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
THE CONSTRUCTION AND USE OF PROMOTER PROBE VECTORS FOR RHODOCOCCUS SP. K. J. KAYSER, C.-O. YUN and J. J. KILBANE II Institute of Gas Technology, 1700 S. Mt. Prospect Road, Des Plaines, IL 60018
Abstract. Three promoter probe vectors have been constructed for use within Rhodococcus sp. They are hybrid replicons capable of replicating both in E.coli and Rhodococcus species due to the presence of replication functions derived from pUC19 and the Rhodococcus fascians plasmid pRF29 respectively. Promoter probe vector pRCM1 contains a promoterless gene which encodes a membrane-associated chloramphenicol efflux protein (cmr) derived from Rhodococcus fascians plasmid pRF2, pRCAT3 contains a promoterless chloramphenicol acetyl transferase gene (cat) derived from Tn9, and pEBC26 contains a promoterless beta-galactosidase gene derived from pSVB-gal. Many derivatives of pRCM1 and pRCAT3 receiving inserts that regulated the expression of chloramphenicol resistance in Rhodococcus sp. strain IGTS8 proved to be unstable in E.coli, frequently yielding plasmids containing deletions. This instability was found to be largely associated with these vectors; however, some inserts of Rhodococcus DNA increased and others alleviated this instability. Derivatives of pEBC26 were stable both in Rhodococcus and E.coli and many DNA fragments encoding Rhodococcus promoters were isolated. The size of these promoter-containing DNA fragments ranged from 0.15 to 3 Kb and the level of beta-galactosidase expression in Rhodococcus hosts ranged from 0.1 to 838 Miller units. Promoters from Rhodococcus were not observed to function in E.coli; however, the E.coli rrnB promoter was shown to function weakly in Rhodococcus.
The genus Rhodococcus comprises gram- positive, aerobic, nonsporulating, partially acid-fast actinomycetes which were formerly classified as Nocardia, Mycobacterium, Gordana, or Jensenia sp.eciesor as members of the rhodochrous complex (Goodfellow et al.,1974; Goodfellow et al.,1977). The genus Rhodococcus is remarkable for its range of activities, which include pathogenicity for human, animal, and plant hosts; degradation of hydrocarbon pesticides, herbicides, and halogenated pollutants and the production of surfactants, stereospecific chemicals, enzymes, and other products (Finnerty, 1992). Rhodococcus cultures are frequently encountered in bioremediation applications because of their ability to degrade a broad range of substrates, particularly hydrophobic compounds. Alkane, cyclic, and aromatic hydrocarbon pollutants can be degraded, and some Rhodococcus cultures possess the rare ability to reductively dehalogenate chlorinated aromatic compounds (Apajalahti & Salkinaja-Salonen, 1986). Rhodococcus cultures also have been used to perform novel biotransformations. Some Rhodococcus cultures have the ability to selectively cleave carbon-sulfur bonds (Kilbane & Jackowski, 1992; Kayser et al., 1993). This activity can be used to desulfurize fossil fuels and as a tool to degrade or modify organosulfur compounds. Other biotransformations accomplished by Rhodococcus cultures include the hydrolysis of complex lipids, the stereospecific oxidation of steroids, the production of acrylamide, and the production of L-phenylalanine (Lechevalier & Lechevalier, 1985).
Despite the widespread interest in Rhodococcus cultures for a variety of applications, there is very little known about the genetics of the genus and next to nothing is known about the control of gene expression in Rhodococcus(Brownell and Deniston, 1984; Finnerty, 1992). Conjugal plasmids capable of chromosomal mobilization have been described and partial genetic linkage maps have been published for R. rhodochrous, R. erythropolis, and R. canicruria.(Finnerty, 1992). Plasmids which encode genes that confer resistance to antibiotics or metals are known for several Rhodococcus species, and some Rhodococcus plasmids are novel conjugal linear plasmids (Apajalahti and Salkinaja-Salonen, 1986; Crespi et al., 1992) Utilizing replication genes and selectable markers from Rhodococcus plasmids and from other microorganisms cloning vectors and shuttle vectors capable of replicating both in E.coli and in Rhodococcus species have been constructed (Singer and Finnerty, 198892; Desomer et al., 1990; Shao et al., 1995). Additionally bacterial viruses for Rhodococcus species have been described including a generalized transducing phage (Brownell and Deniston, 1984). However, cloning vectors or plasmids using Rhodococcus bacteriophages have been discussed, but have not been employed (Finnerty,1992).
Techniques for the chemical transformation of protoplasted Rhodococcus cells and electroporation techniques have been developed and used to introduce plasmid vectors and cloned DNA into a variety of Rhodococcus species (Singer and Finnerty, 1988; Desomer et al., 1990; Shao et al., 1995). However, the majority of genetic analyses that have occurred have involved the cloning of Rhodococcus genes into E.coli (Heiss et al., 1992; Finnerty, 1992; Piddington et al., 1995; and Vereeke et al., 1994). The subsequent analysis of these genes in E.coli has generally been restricted to the determination of the DNA sequences of structural genes of interest. Numerous DNA fragments that most probably encode promoters have been sequenced in the course of characterizing structural genes of Rhodococcus origin, but studies of the expression of Rhodococcus genes are rare (Finnerty, 1992). In general it appears that genes from Rhodococcus are not expressed in E.coli and vice versa and it is clear that the essential features/sequences of Rhodococcus promoters differ from promoters of well characterized microorganisms such as E.coli or B.subtilis (Desomer et al., 1992; Finnerty, 1992).
The gene encoding chloramphenicol resistance (cmr) from the R.fascians plasmid pRF2 may be the sole example of the study of gene expression in Rhodococcus. The promoter of the cmr gene was localized by cloning DNA fragments upstream of a promoterless reporter gene, primer extension experiments were performed to determine the transcriptional initiation site, the DNA sequence was determined, and expression/induction was examined (Desomer et al., 1992). Promoter probe vectors were not available so this study of the cmr promoter was accomplished by the relatively laborious procedure of cloning Rhodococcus DNA fragments suspected of encoding promoters into a B.subtilis-E.coli promoter-probe vector, subcloning an appropriate DNA fragment from these constructs into a vector capable of integrating into the R.fascians chromosome, and finally transforming R.fascians.
The aim of this study was to construct vectors to enable the isolation of DNA fragments from Rhodococcus cultures that encode promoters and to initiate a characterization of Rhodococcus promoters by assessing their ability to function in E.coli hosts.
Materials and Methods
Bacterial Strains, and Culture Conditions. Rhodococcus sp. strain IGTS8 (ATCC No. 53968, Rockville, MD) was used as the source of chromosomal DNA and as the Rhodococcus host strain for genetic experiments. E.coli DH5(Gibco BRL) was used as a host strain for gene cloning and plasmid construction. Rhodococcus sp. strain IGTS8 was cultured at 30^o C in nutrient broth (Difco, Detroit, MI). This medium was supplemented with 1.5% agar (w/v) when solid medium was required and chloramphenicol was used at 25 (g/ml except as indicated otherwise. Recombinant E.coli cells were cultivated at 37^o C in Luria-Bertani (LB) medium (1% tryptone, 0.5% yeast extract, 1% NaCl, pH7.5) containing 100 (g of ampicillin per ml. To detect E.coli or Rhodococcus cells expressing beta-galactosidase activity, 0.01% (w/v) 5-bromo-4-chloro-3-indolyl-B-D- galactopyranoside (X-Gal; Stratagene, La Jolla, CA) was added to solid medium.
Plasmids. Plasmids pGR71 and pUC19 were obtained from ATTC (No. 37105 and No. 37254 respectively). Plasmid pRF29 (Desomer et al., 1990) a derivative of the conjugal Rhodococcus fascians plasmid pRF2 (Desomer et al., 1988), was a gift from Dr. Jan Desomer. Plasmid pKK232-8 was purchased from Pharmacia (Piscataway, NJ), and pSVB-gal was purchased from Promega (Madison, WI). The plasmid pBR16S, which contains the promoter region of the E.coli rrnB operon cloned as a 1.6 Kb BamH1-HindIII fragment cloned into pBR322, was a gift from Dr. Robert Setterquist. Restriction enzymes were purchased from Stratagene (La Jolla, CA) and used according to the suppliers recommendations. Transformation of Rhodococcus was done according to Desomer et al., 1990.
beta - Galactosidase Assay. The method of Miller was used except cells were sonicated rather than treated with sodium dodecyl sulfate. Protein determinations were performed on samples of the same sonicated samples used for determining beta-galactosidase activities, using the kit supplied by Pierce Biochemicals (Rockford, IL). Activities are expressed in Miller units per milligram of protein rather than in units per O.D. 600 unit, since cell size and therefore light-scattering measurements vary slightly at different growth rates.
Results
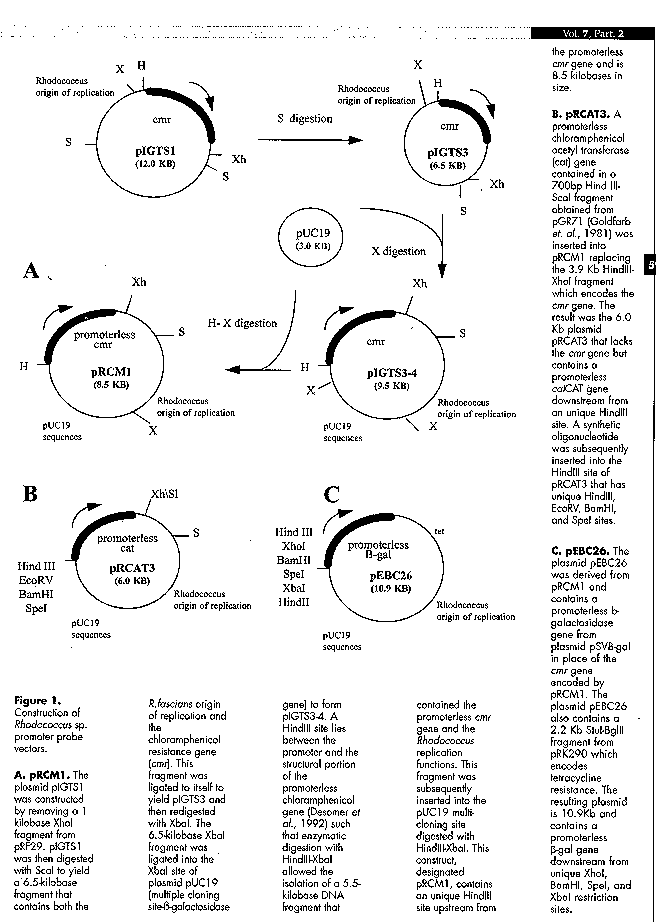
Figure 1A shows the construction of the promoter probe vector pRCM1, which contains the promoterless cmr gene derived from pRF29 that encodes a membrane-associated chloramphenicol efflux protein (Desomer et al., 1992). pRCM1 also contains pRF29 DNA sequences that allow replication in Rhodococcus sp. (Desomer et al., 1992) and it contains pUC19 which allows replication and ampicillin resistance in E.coli. This construct, designated pRCM1, contains an unique HindIII site upstream from the promoterless cmr gene and is 9.2 Kb.
Figure 1B shows pRCAT3, which is derived from pRCM1 by replacement of the cmr reporter gene with a cat reporter gene. The promoter probe vector pRCAT3 contains unique EcoRI, EcoRV, BamHI, and SpeI sites upstream from the promoterless chloramphenicol acetyl transferase gene (cat) derived from pGR71 (Goldfarb et al., 1981) (originally derived from Tn9). pRCAT3 is 6.0 Kb and contains DNA derived from pRF29 that allows replication in Rhodococcus species and pUC19 which allows replication and ampicillin resistance in E.coli.
Figure 1C illustrates pEBC26, which is derived from pRCM1 by the replacement of the cmr reporter gene with the promoterless beta-galactosidase gene from pSVB-gal. The hybrid replicon promoter probe vector pEBC26 is 10.9 Kb and contains unique XhoI, BamHI, SpeI, and XbaI restriction sites upstream from the promoterless b-gal gene. pEBC26 was constructed in collaboration with scientists from Energy BioSystems Corporation (The Woodlands, TX). In addition to containing a promoterless beta-galactosidase gene as a reporter gene, pEBC26 also contains pRF29 sequences which allow replication in Rhodococcus, pUC19 DNA which allows replication and ampicillin resistance in E.coli, and pRK290 sequences which allow tetracycline resistance in both E.coli and Rhodococcus hosts.
Both vectors pRCM1 and pRCAT3 allow the detection of inserted DNA fragments that encode promoters via the chloramphenicol resistance conferred to Rhodococcus transformants. Plasmids pRCM1and pRCAT3 were digested with unique restriction enzymes upstream from the chloramphenicol genes (cmr and cat), then random chromosomal fragments from Rhodococcus sp. strain IGTS8 were ligated into these promoter probe vectors. Numerous chloramphenicol-resistant Rhodococcus sp. strain IGTS8 cultures were isolated from the subsequent transformation of these ligations. Clones containing stable inserts ranging from 100 bp to 2.0 Kb that conferred resistance in Rhodococcus sp. strain IGTS8 to chloramphenicol concentrations ranging from 50 to over 500 æg/ml were obtained. pRCM1 and pRCAT3 plasmid DNA not receiving an insert did not confer chloramphenicol resistance to Rhodococcus or E.coli hosts.
The yield of plasmid DNA from Rhodococcus hosts is low in comparison with yields of the same plasmids from E.coli hosts. This is due to a lower copy number of the plasmids in Rhodococcus and the fact that Rhodococcus is difficult to lyse. Accordingly, plasmid DNA was isolated from chloramphenicol-resistant pRCM1 and pRCAT3-containing Rhodococcus colonies and used to obtain ampicillin resistant E.coli transformants. However, many derivatives of pRCM1 and pRCAT3 that express chloramphenicol resistance in Rhodococcus sp. strain IGTS8 proved to be unstable in E.coli, frequently yielding plasmids containing deletions. Moreover, some plasmid DNA samples could not be used to obtain transformants in E.coli. The instability of derivatives of pRCM1 and pRCAT3 containing inserts of Rhodococcus DNA, in E.coli, was investigated further.
A. pRCM1. The plasmid pIGTS1 was constructed by removing a 1 kilobase XhoI fragment from pRF29. pIGTS1 was then digested with ScaI to yield a 6.5-kilobase fragment that contains both the R.fascians origin of replication and the chloramphenicol resistance gene (cmr). This fragment was ligated to itself to yield pIGTS3 and then redigested with XbaI. The 6.5-kilobase XbaI fragment was ligated into the XbaI site of plasmid pUC19 (multiple cloning site-beta- galactosidase gene) to form pIGTS3-4. A HindIII site lies between the promoter and the structural portion of the promoterless chloramphenicol gene (Desomer et al., 1992) such that enzymatic digestion with HindIII-XbaI allowed the isolation of a 5.5-kilobase DNA fragment that contained the promoterless cmr gene and the Rhodococcus replication functions. This fragment was subsequently inserted into the pUC19 multi-cloning site digested with HindIII-XbaI. This construct, designated pRCM1, contains an unique HindIII site upstream from the promoterless cmr gene and is 8.5 kilobases in size. B. pRCAT3. A promoterless chloramphenicol acetyl transferase (cat) gene contained in a 700bp Hind III- ScaI fragment obtained from pGR71 (Goldfarb et. al., 1981) was inserted into pRCM1 replacing the 3.9 Kb HindIII- XhoI fragment which encodes the cmr gene. The result was the 6.0 Kb plasmid pRCAT3 that lacks the cmr gene but contains a promoterless catCAT gene downstream from an unique HindIII site. A synthetic oligonucleotide was subsequently inserted into the HindIII site of pRCAT3 that has unique HindIII, EcoRV, BamHI, and SpeI sites. C. pEBC26. The plasmid pEBC26 was derived from pRCM1 and contains a promoterless beta-galactosidase gene from plasmid pSVB-gal in place of the cmr gene encoded by pRCM1. The plasmid pEBC26 also contains a 2.2 Kb StuI-BglII fragment from pRK290 which encodes tetracycline resistance. The resulting plasmid is 10.9Kb and contains a promoterless b- gal gene downstream from unique XhoI, BamHI, SpeI, and XbaI restriction sites.
The native promoter of the cmr gene, contained on a 700 bp HindIII-XbaI fragment from pRF29, was cloned into the HindIII-SpeI sites of pRCAT3 upstream from the promoterless cat gene. Chloramphenicol-resistant transformants in Rhodococcus were only observed when this promoter fragment was inserted in pRCAT3 in the same orientation relative to the cat gene as exists in the native cmr gene. DNA isolated from chloramphenicol resistant Rhodococcus colonies containing these specific derivatives of pRCAT3 yielded ampicillin resistant transformants in E.coli at low frequency and these transformants often contained obvious deletions. Chloramphenicol resistant E.coli transformants were not obtained.
A 1.6 Kb BamH1/HindII fragment containing the promoter from the rrnB operon of E.coli was obtained from pBR16S and was ligated into pRCAT3 DNA that had been cleaved with BamH1 and HindIII. The resulting plasmid, pE16CAT, yielded E.coli transformants that were resistant to over 1000 ug/ml chloramphenicol; however, chloramphenicol resistant Rhodococcus transformants could not be obtained despite repeated attempts.
Rhodococcus chromosomal fragments were inserted into the E.coli promoter probe vector pKK232-8, which has a cat reporter cassette. Numerous inserts ranging from 100 bp to more than 2 Kb that conferred chloramphenicol resistance in E.coli were obtained. These derivatives of pKK232-8 allow the growth of E.coli colonies on agar plates containing from 25 to 200 micrograms chloramphenicol per ml, and no instability/deletions was observed.
To further identify DNA fragments from Rhodococcus that encode promoters genomic DNA from Rhodococcus sp. strain IGTS8 was digested with Sau3A and cloned into the BamH1 site of pEBC26 to create a Rhodococcus promoter library. Hundreds of transformants were obtained in Rhodococcus that expressed various levels of beta- galactosidase. DNA from each beta-galactosidase-positive Rhodococcus colony uniformly yielded stable ampicillin resistant transformants in E.coli. These E.coli transformants did not express beta-galactosidase at levels significantly higher than control cultures containing pEBC26 only. The 1.6 Kb BamH1-/HindIII fragment of pBR16S, which contains the promoter region of the E.coli rrnB operon, was cloned into pEBC26 yielding pE16PRO. When this plasmid was transformed into Rhodococcus stable tetracycline resistant derivatives were obtained that expressed beta- galactosidase at 12 units. However, stable E.coli transformants could not be obtained and ampicillin resistant colonies were uniformly observed to generate multi-sectored colonies comprised of beta-galactosidase positive and beta- galactosidase negative sectors. Subsequent analysis of beta- galactosidase negative sectors revealed that they contained ampicillin sensitive cells, and no plasmid DNA could be detected. Derivatives of pEBC26 that contain DNA fragments which encode Rhodococcus promoters did not give rise to sectored E.coli colonies. Table 1 summarizes the preliminary characterization of some pEBC26 derivatives that contain promoters derived from Rhodococcus DNA.
-------------------------------------------------------------- Table 1. Preliminary characterization of Rhodococcus promoters isolated using the beta-galactosidase promoter probe vector pEBC26. The size of chromosomal DNA fragments in each clone as determined by restriction endonuclease analysis and the specific beta-galactosidase activity in each clone in a Rhodococcus host as determined according to Miller (1972) is indicated. -------------------------------------------------------------- Plasmid Insert Size (Kb) Specific Activity (beta-gal) -------------------------------------------------------------- pEBC26 None 0 pYGALa 1.2 838 pYBALb 0.4 50 pYGALc 0.8 52 pYGALd 3.0 23 pYGALe 0.2 49 pYGALf 1.1 53 pYGALg 2.3 19 pYGALh 1.1 44 pYGALi 0.3 111 pYGALj 0.6 131 pYGALk 0.3 104 pYGALk2 0.3 71 pYGALl 0.15 1.4 pYGALm 0.15 31 pYGAL13 2.2 0.1 pYGAL14 0.6 140 pYGAL15 0.8 72 pYGAL16 1.3 500 pYGAL17 0.5 142 pYGAL18 0.6 26 pYGAL19 1.4 48 pYGAL20 1.6 26 pYGAL21 0.4 58 pYGAL22 0.6 66 pYGAL23 0.4 21 pE16PRO 1.6 12 -------------------------------------------------------------- Discussion
Isolation of plasmid DNA from Rhodococcus is relatively difficult because Rhodococcus cells are often recalcitrant to common lysis and plasmid-isolation protocols, and the copy number for pRF29 and related plasmids in Rhodococcus is low (Desomer, 1990). The low yields of plasmid DNA from Rhodococcus hosts renders DNA manipulation in E.coli far more convenient than in Rhodococcus. Thus, promoter probe vectors were constructed that replicate in Rhodococcus as well as in E.coli.
Numerous Rhodococcus chromosomal fragments cloned into pRCM1 and pRCAT3 yielded chloramphenicol resistant Rhodococcus sp. strain IGTS8 transformants. Although these insert-containing derivatives of pRCM1 and pRCAT3 were stable in Rhodococcus they frequently were found to be unstable in E.coli. The promoter probe vectors pRCM1 and pRCAT3 were themselves unstable in E.coli, and this instability was exacerbated, alleviated, or unaffected in both vectors in E.coli when they contained chromosomal DNA inserts from Rhodococcus. Since these vectors do not contain selectable markers expressed in Rhodococcus their stability in Rhodococcus cannot be tested directly. The fact that stable chloramphenicol resistant derivatives of pRCM1 and pRCAT3 are obtained in Rhodococcus could indicate that the instability only occurs in E.coli hosts; however, the act of selecting chloramphenicol resistant Rhodococcus colonies could bias the results allowing only stable derivatives to be isolated. Some derivative plasmids are also stable in E.coli, but this is observed far less frequently. The cause of the instability of these vectors in E.coli remains unknown, and this is particularly puzzling since the E.coli host used, DH5a, is recA-negative. An explanation is also lacking for the reason that some Rhodococcus DNA fragments stabilize these vectors and some DNA fragments aggravate the instability of these vectors. E.coli transformants exhibiting chloramphenicol resistance were not observed with either pRCM1 or pRCAT3 derivatives further supporting the hypothesis that Rhodococcus promoter sequences do not function in E.coli. When the E.coli reporter vector pKK232-8 had Rhodococcus chromosomal fragments inserted upstream from the cat cassette, chloramphenicol resistance was conferred in E.coli, and the constructs were stable. Niether increased nor decreased stability of these plasmids relative to pKK232-8 was observed. Since pKK232-8 cannot replicate in Rhodococcus it was not possible to ascertain if the Rhodococcus DNA fragments that functioned as promoters in E.coli in these pKK232-8 derivatives were authentic Rhodococcus promoters, but in any event the addition of Rhodococcus DNA per se does not seem to affect the stability of this E.coli plasmid. While this work was in progress a promoter probe vector, pDP1, was reported that utilizes the pRF2 origin of replication, the cmr gene as a promoterless reporter gene, and pUC19 for replication and ampicillin resistance in E.coli. Thus pDP1 (Vereeke, 1994) is highly similar to pRCM1; however, instability was not reported associated with the use of pDP1.
The promoter probe vectors pRCM1 and pRCAT3 were successfully used to isolate Rhodococcus DNA fragments that encode promoters and a collection of fragments conferring resistance to 50 to over 500æg chloramphenicol/ml were obtained. However the unstable nature of these vectors limits their use so an alternative promoter probe vector was constructed.
The results obtained with pEBC26 and illustrated in Table 1 demonstrate that authentic Rhodococcus promoters can be isolated using this promoter probe vector with relative activities that differ by as much as about 800 fold. Furthermore the plasmids listed in Table 1 can be stably maintained in E.coli. However, the promoters contained in these pEBC26 derivatives do not appear to function in E.coli resulting in higher levels of beta-galactosidase expression, but this conclusion is qualified by the fact that pEBC26 exhibits a significant background level of beta- galactosidase expression in E.coli (data not shown).
Tools for the isolation and characterization of Rhodococcus DNA involved in the regulation of genetic expression have been developed. Since pRF29 replicates and expresses chloramphenicol resistance in many different Rhodococcus species, (Bielaga, 1991) promoter probe vectors derived from pRF29 such as pEBC26 and pDP1 should prove to be useful across the genus. Multiple DNA fragments encoding functional Rhodococcus promoters of various strengths have been isolated. None of these Rhodococcus promoters was observed to function in E.coli. The promoter of the rrnB operon of E.coli was shown to function weakly in Rhodococcus, but would contribute to plasmid instability when cloned in E.coli. Instability of cloned ribosomal promoters has been observed previously and results from the metabolic strain placed on cells containing cloned versions of these strong rrn operon promoters. This instability of rrn promoter- containing plasmids is most probably different from the nature of the instability of pRCM1 and pRCAT3 in E.coli. Since rrn operons are among the most highly conserved genes known, and rrn promoters are among the strongest promoters known, the fact that an E.coli rrn promoter was only capable of functioning weakly in Rhodococcus suggests that the factors governing genetic expression in E.coli and Rhodococcus are quite diverse, perhaps suprisingly so. DNA sequence analyses of Rhodococcus promoters and transcriptional analysis of Rhodococcus promoters both in E.coli and in Rhodococcus is currently in progress.
Acknowledgments. This work was prepared with the support of Energy BioSystems Corporation and in part by grants made possible by the Illinois Department of Energy and Natural Resources through its Coal Development Board and Illinois Clean Coal Institute, and by the U.S. Department of Energy (DE-FC22-92PC92521). However, any opinions, findings, conclusions, or recommendations expressed herein are those of the author and do not necessarily reflect the views of IDENR, ICCI, and the DOE. The technical assistance of Chris McDonald and Karen Lawrence is gratefully acknowledged.
References Apajalahti, J.H.A. & M.S.Salkinaja-Salonen (1986). Degradation of polychlorinated phenols by Rhodococcus chlorophenolicus. Appl. Microbiol. Biotechnol., 25: 62-67 Bielaga, B.A. (1991). Genetic analysis of Rhodococcus rhodochrous IGTS8. Masters Thesis, Illinois Institute of Technology, Chicago, Illinois Brownell, G.H. & K.Deniston (1984).Genetics of nocardioform bacteria. In: M.Goodfellow, M.Mordarski & S.T.Williams (eds.) The Biology of Actinomycetes. Academic Press, New York, pp. 201-208 Crespi, M., E.Messens, A.B.Caplan, M.Van Motagu & J.Desomer (1992). Fasciation induction by the phytopathogene Rhodococcus fascians depends upon a linear plasmid encoding a cytokinin synthase gene. EMBO J., 11: 795-804 Desomer, J., P.Dhase & M.Van Montagu (1988). Conjugative transfer of cadmium resistance plasmids in Rhodococcus fascians strains. J. Bacteriol., 170: 2401- 2405 Desomer, J., P.Dhaese & M.Van Montagu (1990). Transformation of Rhodococcus fascians by high-voltage electroporation and development of R.fascians cloning vectors. Appl. Environ. Microbiol., 56: 2818-2825 Desomer, J., D.Vereecke, M.Crespi & M.Van Montagu (1992). The plasmid-encoded chloramphenicol-resistance protein of Rhodococcus fascians is homologous to the transmembrane tetracycline efflux proteins. Molec. Microbiol., 6: 2377-2385 Finnerty, W.R. (1992).The Biology and Genetics of the Genus Rhodococcus. Ann. Rev. Microbiol., 46: 193-218 Goldfarb, D.S., R.Doi & R.L.Rodriguez (1981). Expression of Tn9-derived chloramphenicol resistance in Bacillus subtillis. Nature, 293: 309-311 Goodfellow, M., A.Lind, H.Mordarska, S.Pattyn & M.Tsukamura (1974). A cooperative numerical analysis of cultures considered to belong to the 'rhodochrous' taxon. J. gen. Microbiol., 85: 291-302 Goodfellow, M. & G.Alderson (1977). The actinomycete-genus Rhodococcus: A home for the 'rhodochrous' complex. J. gen. Microbiol., 100: 99-122 Heiss, G.S., B.Gowan & E.R.Dabbs (1992). Cloning of DNA from a Rhodococcus strain conferring the ability to decolorize sulfonated azo dyes. FEMS Microbiol. Lett., 99: 221-226 Kayser, K.J., B.A.Bielaga-Jones, K.Jackowski, O.Odusan & J.J.Kilbane (1993).Utilization of organosulfur compounds by axenic and mixed cultures of Rhodococcus rhodochrous IGTS8. J. gen. Microbiol., 139: 3123-3129 Kilbane, J.J. (1989). Desulfurization of coal: The microbial solution. Trends in Biotechnology, 7 (4): 97- 101 Kilbane, J.J. & K.Jackowski (1992). Biodesulfurization of water-soluble coal-derived material by Rhodococcus rhodochrous IGTS8. Biotechnol. Bioeng., 403: 1107-1114 Lechevalier, M.P. & H.Lechevalier (1985). Biology of actinomycetes not belonging to genus Streptomyces. In: A.I.Demain & N.A.Solomon (eds.) Biology of Industrial Microorganisms. Benjamin/Cummings, Menlo Park, CA., pp. 315-358 Miller, J.H. (1972). Experiments in Molecular Genetics. Cold Springs Harbor Laboratory, Cold Spring, New York, pp. 352-355 Piddington, C.S., B.R.Kovaceich & J.Rambosek (1995). Sequence and molecular characterization of a DNA region encoding the dibenzothiophene desulfurization operon of Rhodococcus sp. strain IGTS8. Appl.. Environ. Microbiol., 61: 468-475 Shao, Z., W.A.Dick & R.M.Behki (1995). An improved Escherichia coli-Rhodococcus shuttle vector and plasmid transformation in Rhodococcus spp. using electroporation. Lett. Appl. Microbiol., 21: 261-266 Singer, M.E. & W.R.Finnerty (1988). Construction of an Escherichia coli-Rhodococcus shuttle vector and plasmid transformation in Rhodococcus sp. J. Bacteriol., 170: 638-645. Copyright 1996 C.E.T.A., The International Centre for Theoretical and Applied Ecology, Gorizia The following images related to this document are available:Line drawing images[ac96009a.gif] |
| |||||||||
{kind=link}