 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
DEGRADATION OF COMPLEX MINERAL OIL AND OTHER HYDROCARBONS BY RHODOCOCCUS ERYTHROPOLIS I. SAADOUN Department of Biological Sciences, Jordan University of Science and Technology, Irbid-22110, Jordan
Code Number: AC97009
Sizes of Files:
Text: 9.7K
Graphics: Line drawing (gif) - 5K
ABSTRACT. The ability of Rhodococcus erythropolis to degrade mineral oil and other hydrocarbons namely, hexane, decane and tetradecane was determined by using a Warburg constant volume respirometer. Results of oxygen uptake indicated that hexane and tetradecane were more degradable than mineral oil and decane. R. erythropolis exhibited highest QO2 values (2.859 and 2.816) when exposed to tetradecane and hexane, respectively. Mineral oil and decane were degraded more slowly with QO2 values 0.866 and 0.939 respectively. Petroleum pipeline ruptures, tank failures and other production, storage and transportation accidents contribute to soil hydrocarbon contamination (Bossert et al. 1984). Accidental contamination of this type is classified as hazardous waste (Barth et al. 1984).
Hydrocarbon degradation in soil by microorganisms (Dagley, 1975) has undergone renewed emphasis because of the increased incidence of petroleum-based pollution. Knowledge about hydrocarbon degradation is needed to determine how microorganisms might be utilised in the removal of the pollutants from the environment. An actinomycete, isolated from a commercial bacterial consortium (Blevins et al,. 1993), and identified as Rhodococcus erythropolis has been used to study the degradation of different hydrocarbons by substrate reduction as determined by oxygen uptake in respiratory processes. The objective of the present study is to investigate, by oxidation studies, the ability of the organism to degrade different hydrocarbon substrates. MATERIALS and METHODS Organism. Rhodococcus erythropolis was identified by fatty acid methyl ester analysis (MIDI) identification system (Blevins et al., 1993).
Adaptation of the degrader. R. erythropolis was incubated overnight in 50 ml yeast-dextrose broth (yeast extract, 10g; 10g glucose, 10g; pH 7.5) at 28 C (shaken culture: 200rpm). Cells were centrifuged and washed twice with the liquid inorganic salts medium (MSM) of Leadbetter and Foster (1958). The pellet was suspended in 5ml MSM and inoculated into 500ml of MSM in 21 flasks (supplemented with 0.001% yeast extract).
Growth on mineral oil was determined by measuring the optical density (O.D.) at 615 nm with a Spectronic Colorimeter 20 (Bausch and Lomb Inc., Rochester, NY). Wet mounts were examined by a phase-contrast microscope (Olympus Optical Co., Ltd., Tokyo, Japan) and a Nikon autoexposure unit. Mineral oil adapted cells were harvested and washed twice with MSM and the pellet suspended in 0.1M phosphate buffer at pH 7.0.
Biodegradation of hydrocarbons. Determination of total oxygen consumption by bacterial ceils was carried out with a Warburg constant volume respirometer (Precision Scientific Co., Chicago, Ill., USA). The procedure followed was that suggested by Umbreit et al. (1964). Single sidearm flasks with a centrally located KOH cup were used for setting up the experiment.
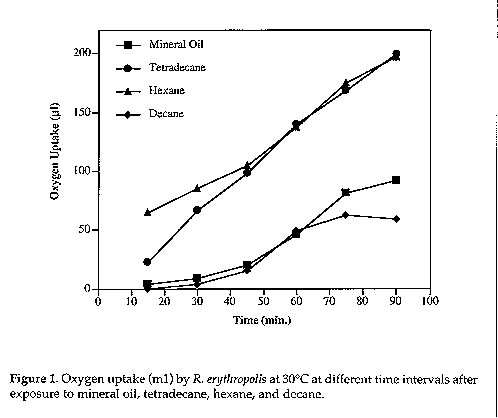
Thermobar flasks were prepared by adding 2.2ml of 0.1M phosphate buffer in the main chamber, 0.2ml of 0.1M phosphate buffer were placed in the side arm and 2ml of cells suspended in 0.1M phosphate buffer in the main chamber. Test flasks were prepared as follows: 0.2ml of the substrate were placed in the sidearm, 2ml of the cell suspension in the main chamber, 0.2ml of 20% KOH and fluted filter paper were placed in the centrally located cup of each flask. Flasks were kept in a water bath (30 C) with 10 min shaking. The level of Brodie's solution (23g NaCl, 5g Na-taurocholate dissolved in 500ml water, one drop of alcoholic thymol and acid fuchsin) in the right side of each manometer was adjusted to 150mm, then the stop-cocks were closed and the left side level of the manometer was recorded at zero time. Meanwhile, the substrate in the sidearm was tilted to mix it with the cell suspension in the main chamber. Oxygen uptake was followed at 30 C for 1.5hr with readings being taken at 15 min intervals. Dry weight determinations were carried out on 2ml of the cell suspensions dried at 60 C for 48hrs. RESULTS and DISCUSSION Adaptation of R. erythropolis to mineral oil and ability to degrade complex hydrocarbons was preliminary judged by turbidity. Growth was determined spectrophotometrically (Table 1) and results indicate the ability of the organism to utilise the substrates as sole carbon sources. Hexane and tetradecane were degraded more effectively than mineral oil and decane. Table 1. Respiratory quotient of R. erythropolis exposed to different hydrocarbons and glucose. Values are averages of three replicates, corrected for endogenous rates.
----------------------------
SUBSTRATE Q02
----------------------------
Mineral oil 0.866
Hexane 2.816
Decane 0.939
Tetradecane 2.859
Glucose 0.448
----------------------------
Growth on mineral oil before exposure to specific hydrocarbon compounds was intended as a pre-enrichment step, similar to the initial enrichment of polychlorinated biphenyl (PCB) degraders on biphenyl (Bedard et al., 1987) or methylcyclohexane for bacteria able to grow on a wide range of alicyclic compounds (Trudgill, 1984).
Hydrocarbons that are resistant to both oxidation and assimilation when present alone in the medium, may be oxidised if attacked by organisms that are simultaneously oxidising other hydrocarbons. This process, called cooxidation, was noted by Leadbetter & Foster (1958) when investigating the oxidation of ethane, propane, and butane by Pseudomonas mechanica while growing on methane, the only hydrocarbon supporting growth. Growth of R. erythropolis on complex mineral oil was required to incite the decomposition of the tested hydrocarbons.
There is no evidence that all of the different hydrocarbons are decomposed by this bacteria by co-metabolism or co-oxidation metabolism.
In this study, results indicate that tetradecane and hexane are more easily degradable than mineral oil and decane as evidenced by the higher oxygen uptake (Fig. 1). Degradation of tetradecane and hexane is demonstrated by oxygen uptake (2.859 and 2.816ml of oxygen/mg cells in 60 min, respectively). Saadoun (1995) reported lower values (0.09 and 0.18) for R. erythropolis when exposed to cyclohexane and toluene respectively. QO2 values obtained in this study can be of use in the comparison of microbial oxidation of various hydrocarbon compounds.
REFERENCES Barth, R. & I. Bossert (1984). The treatment and disposal of petroleum refinery wastes, In: R.M. Atlas (ed.) Petroleum Microbiology. Macmillan, New York, pp. 553-577 Bedard, D.L., R.E. Wagner, M.J. Brennan, M.L. Harbel & J.F. Brown, Jr (1987). Extensive degradation of arochlors and environmentally transformed polychlorinated biphenyl's by Alcaligenes eutrophus H850. Appl. Environ. Microbiol., 53:1094-1102 Blevins, W.T., W.T. Crutchfield, K.K. Schrader & I. Saadoun (1993). Species diversification of a microbial consortium during biodegradation of a complex oil. Abstr. Am. Soc. Microbiol., p.361 Bossert, I. & R. Barth (1984). The fate of petroleum in soil ecosystems. In: R.M. Atlas (ed.) Petroleum Microbiology. Macmillan, New York, pp. 453-473 Dagley, S. (1975). A biochemical approach to some problems of environmental pollution. Essays Biochem. 11:81-138 Leadbetter, E.R. & J.W. Foster (1958). Studies of some methane utilizing bacteria. Arch. Microbiol. 30:91-118 Saadoun, I. (1995). Physiology and genetics of off favor compound production by aquatic microorganisms. Ph.D. Dissertation, Department of Botany and Microbiology, Auburn University, Auburn, Alabama. Trudgill, P.W. (1984). Microbial degradation of the alicyclic ring. In D.T. Gibson (ed.), Microbial degradation of organic compounds. Marcel, Dekker, Inc., New York Umbreit, W.W., R.H. Borris & J.F. Stauffer (1964). Manometric Techniques. Burgess Publ. Comp., Minneapolis, Minnesota pp. 305. Copyright 1997 C.E.T.A., The International Centre for Theoretical and Applied Ecology, Gorizia The following images related to this document are available:Line drawing images[ac97009a.gif] |
| |||||||||
{kind=link}