 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
DETECTION OF GIANT LINEAR PLASMIDS IN OFF-FLAVOR COMPOUND-PRODUCING STRAINS OF STREPTOMYCES BY PFGE I. SAADOUN and W.T. BLEVINS^1
Department of Biological Sciences, Jordan University of Science and
Technology, Irbid-22110, Jordan and
Code Number: AC97010
Sizes of Files:
Text: 21.2K
Graphics: Photographs (jpg) - 100.2K
ABSTRACT. A geosmin-producing actinomycete, identified as Streptomyces halstedii and isolated from a catfish production pond, and a MIB-producer, isolated from a drinking water reservoir and identified as S. violaceusniger, were used to investigate the presence of plasmids. Several lytic techniques were compared for recovery of chromosomnal and plasmid DNA. Using an alkaline-lysis DNA extraction procedure and conventional gel electrophoresis, no low molecular weight plasmids were observed in any of the isolates tested. However, lysis of Streptomyces cells in low melting point agarose followed by pulsed-field electrophoresis (CHEF-DR II) of the DNA yielded high-molecular weight plasmids. Various species of actinomycetes and cyanobacteria can impart earthy/musty off-flavors to drinking water supplies and to pond-raised fish and other aquatic food animals. Geosmin (trans-1,10-dimethyl-trans-9-deca-1ol) and 2-methylisoborneol (1,2,7,7-tetramethyl-exo-bicyclo-[2,2,1]-heptan-2ol or MIB) are secondary metabolites produced by cyanobacteria (Tabachek & Yurkowski, 1976; Izaguirre et al., 1982) as well as actinomycetes (Gerber, 1979; Bentley & Meganathan, 1981) and these compounds can impart earthy/musty off-flavors (or tastes and odors) to the aquatic environment.
There are few studies regarding the genetic basis for the biosynthesis of these off-flavor compounds or the regulation of their synthesis. Some reports suggest that the genetic determinants for off-flavor compounds in actinomycetes may be carried on plasmids (Redshaw et al., 1979; Ishibashi, 1992). Plasmids are abundant in the genus Streptomyces, and usually exist as covalently closed circular (CCC) DNA molecules. Some species harbor linear DNA plasmids, with streptomycetes being the only prokaryotic organisms reported to contain linear plasmids (Hayakawa et al., 1979; Hirochika & Sakaguchi, 1982; Hirochika et al., 1984; Chadron-Loriaux et al., 1986; Kinashi & Shimaji, 1987; Keen et al., 1988; Ishibashi, 1992).
Pulsed-field gel electrophoresis (PFGE) has been used to detect a giant (500 kb) linear plasmid proposed to encode for geosmin production in Streptomyces lasaliensis (Ishibashi, 1992). Treatment of Streptomyces species with ethidium bromide (EB), acriflavine (AF), or acridine orange (AO) can generate "bald" mutants which have lost the ability to sporulate and may also lack the ability to synthesize off-flavor compounds (Redshaw et al., 1976; Redshaw et al., 1979). Transposable elements, which are also mutagenic, could contribute to loss of odor-producing ability due to chromosomal mutations induced by these agents (Sermonti et al., 1980).
In this study, various plasmid DNA extraction methods were used to extract plasmid DNA from a geosmin-producing actinomycete, identified as S. halstedii, and from a MIB-producing actinomycete, identified as S. violaceusniger MATERIALS and METHODS Microbial cultures. The following microbial cultures were used in this study: 1) Streptomyces halstedii: a geosmin-producing actinomycete isolated from a catfish production pond; 2) S. violaceusniger: a MIB-producing actinomycete isolated from sediment of a local stream; 3) S. lividans ATCC 35287, containing pIJ702 plasmid (5.65 kb in size), and used as a control; and 4) Escherichia coli strain X2556, containing eight plasmids ranging in size from 1.4 to 35 megadalton (Mdal), used as a source of molecular weight standards. Growth conditions. Actinomycetes were cultured on either yeast extractdextrose (YD) agar [10g/l dextrose, 10g/l yeast extract (YE), 0.5% (v/v) glycerine, pH 7.5], or yeast extract-malt extract (YEME) broth (Hopwood et al., 1985) (3g/l YE, 5g/l Bacto-peptone, 3g/l malt extract, 10g/l glucose, 340g/l sucrose, 0.5% glycine and 2ml of 2.5M MgCl2.6H2O; the latter was added after autoclaving). Actinomycetes were grown in broth cultures at 28 C with shaking at 200rpm for 3 days. E. coli cultures were grown at 37 C with shaking at 200rpm overnight.
Recovery of plasmid DNA. The alkaline lysis method of Kieser (1984) was followed for detection of small molecular weight plasmid DNA (covalently closed circular or CCC). S. lividans ATCC 35287, which harbors pIJ702 plasmid of 5.65kb, was used as a positive control. Conventional gel electrophoresis was usually conducted with 0.7% agarose gels using a Tris-borate (O.045M)-EDTA (O.001M) buffer (0.5x TBE). Electrophoresis was carried out at a constant current of 35mA. High molecular weight plasmid DNA from protoplasts of S. halstedii and S. violaceusniger was detected by pulsed-field gel electrophoresis (PFGE) according to the protocol of Kinashi (1987). Electrophoresis was conducted with 1% PFGE agarose (Biorad, Hercules, CA) in 0.5 x TBE at 180V for 18 hr, with a switching time of 30-60 sec at 12 C using a contour-clamped homogeneous electric field (CHEF-DR-II) (Biorad, Hercules, CA) system. All gels were stained with a solution of 0.5mg/ml ethidium bromide for 20 min at room temperature. Gels were destained by rinsing with distilled water and viewed using an ultraviolet (302nm) transilluminator Model TM-36 (Ultra Violet Products Inc., San Gabriel, CA). The gels were then photographed using a Polaroid DS-34 Direct Screen Instant Camera (Polaroid Corporation, Cambridge, MA) fitted with a Tiffen 40.5mm deep yellow 15 filter (Tiffen Manufacturing Co., Hauppauge, NY).
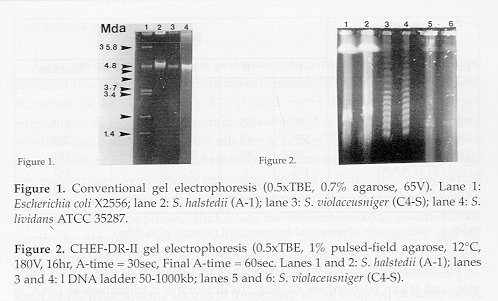
Restriction endonuclease digestion of plasmid DNA. Plasmid DNA bands cut out of agarose gels were kept in 0.5M EDTA (pH 8). Agarose plugs (with DNA) of 1 x 3 x 10 mm size were dialyzed overnight in TEN (10mM Tris-HCl, pH 7.4; 1mM EDTA and 50mM NaCl) buffer, then incubated with 89ml of sterile distilled water, 10ml restriction buffer and 1m of bovine serum albumin (BSA, 1mg/ml) at 37 C for 5 min in 1.5ml Eppendorf tubes. Restriction endonuclease (RE) digestions were carried out according to the instructions of the supplier (Promegg Co., Madison, WI, and Boehringer-Mannheim Biochemica Co., Indianapolis, IN) for complete digestion at 37 C/16hr. The resulting products were separated in 1% agarose gels in 0.5 x TBE at 180V and 12 C for 15 hr with a switching time of 1-45 sec using the CHEF-DR II system. Sizes of DNA fragments were determined by comparing with digests of lDNA as markers (Promegg Co., Madison, WI). Molecular weights and/or sizes of plasmid DNA fragments reported represent averages of several determinations. RESULTS Testing for small CCC plasmid DNA. For the initial experiments to detect plasmids in actinomycetes, S. lividans (ATCC 35287) harboring the 5.65-kb plasmid pIJ702 (Katz, 1983) and a strain of E. coli (X2556) (same as VA 517 according to Macring et al., 1978) were chosen as positive controls and size markers, respectively. No small CCC or large linear plasmid DNA were detected in either aquatic Streptomyces isolates using conventional agarose gel electrophoresis (Fig. 1, lanes 1 and 2, respectively).
Detection of linear plasmid DNA. DNA samples prepared by conventional methods showed no plasmid bands (Fig. 1) for Streptomyces. A more gentle DNA lysis was used by embedding Streptomyces protoplasts in low-melting agarose followed by treatment with pronase and SDS in the gel to prevent physical shearing of DNA (Kinashi et al., 1987), followed by pulsed field gel electrophoresis (CHEF-DR II system); this procedure clearly revealed a plasmid band in both geosmin- and MIB-producers (Fig. 2). The presence of giant linear plasmid DNA of approximately 180kbp (Fig. 2, lanes 1 and 2) in S. halstedii and 115kbp (Fig. 2, lanes 5 and 6) in S. violaceusniger was confirmed.
Figure 2. CHEF-DR-II gel electrophoresis (0.5 x TBE, 1% pulsed-field agarose, 12 C, 18 0V, 16hr, A-time = 30 sec, Final A-time = 60 sec. Lanes 1 and 2: S. halstedii (A-1); lanes 3 and 4:1 DNA ladder 50-1000kb; lanes 5 and 6: S. violaceusniger (C4-S).
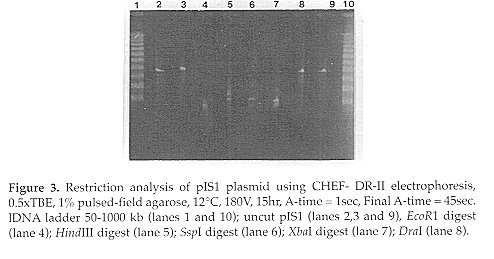
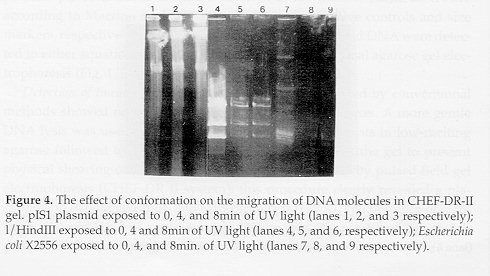
------------------------------------------------------ FRAGMENT I II III IV Total kbp ------------------------------------------------------ EcoRl (G/ AATTC) - - - - - HindIII(A/AGCTT) 95 55 35 25 210 Sspl (AAT/ATT) 126 62 42 - 230 Xbal (T/CTAGA) 82.5 48 33 20.5 184 Dral (TTT/AAA) 182 - - - 182 Uncut pIS1 180 - - - 180 ------------------------------------------------------ The pIS1 plasmid is not circular. Linear DNA is known to migrate in CHEF gels, and it can be resolved by size. To determine if migration of DNA in CHEF gels was affected by a circular conformation, the migration of the supercoiled plasmid DNA of E. coli X2556 and the linear DNA of the 1 / HindIII digest after UV light exposure was determined. As shown in Fig. 4, a majority of the supercoiled plasmid DNA remained in the well, and the brightness of the well area increased with progressive exposure to ultraviolet light (lanes 7, 8 and 9). However, the bands of the linear DNA of the 1/HindIII digest entered the gel and were not affected by UV light exposure. Furthermore, circular DNA moves much more slowly than linear DNA on the CHEF-DR II gel; for example CCC DNA of E. coli X2556 of 54kbp (35.6 Mdal) migrates similarly to linear 6.6kbp DNA fragment produced from 1 by HindIII. Furthermore, pIS1 plasmid DNA remained in the same position relative to 1/HindIII linear DNA under different switching conditions, which clearly shows that it behaves like linear DNA (Fig. 2, 3 and 4).
Many attempts at isolation of plasmids from S. halstedii and S. violaceusniger failed to reveal any covalently closed circular DNA species. The alkaline extraction of DNA (Kieser, 1984) yielded better recovery of DNA from Streptomyces sp. than other methods (Saadoun et al., 1993). This procedure involved incubation of the DNA mixture at higher temperatures that lead to denaturation of linear DNA and hydrolysis of most of the RNA. Furthermore, any method employing phenol extraction can lead to hydrodynamic shearing forces that might cause loss of a linear plasmid at the interface between the aqueous and organic phases. Thus, linear double-stranded (ds) plasmid DNA should be eliminated under the conditions used (Fig. 1, lanes 1 and 2). Application of the CHEF-DRII technique in combination with DNA preparation using gentle lysis enabled the detection of giant linear plasmids in geosmin and MIB-producing strains of Streptomyces. The isolation of linear DNA plasmids from S. halstedii (pISl) and S. violaceusniger (pIS2) was accomplished, and isolation of these plasmids could only be achieved using a gentle extraction procedure that diminishes physical shearing processes.
Restriction endonuclease susceptibility studies suggested that the pIS1 plasmid is linear as digestion resulted in a minimum of two fragments. Furthermore, the size of the fragments was consistent with the apparent length of the uncut plasmid, assuming it is linear; however, the size of pIS1 linear plasmid was over estimated when restricted with HindIII, Sspl, and Xbal. This can be explained by the partial digest of the DNA as evidenced by the faint bands. Further confirmation of the linearity of the pIS1 plasmid was revealed as follows: 1) circular DNA moves much more slowly than linear DNA on the CHEF-DRII gel; for example CCC-DNA of E. coli X2556 of 54kb (35.6Mdal) (1kb = 0.66Mdal) co-migrates like the linear 6.6 kbDNA fragment from 1 digested by HindIII (Fig. 4, lanes 4 and 7); additionally no small circular plasmid band was seen in a conventional agarose gels (Fig. 1) from alkaline preparations of strain A-1; 2) pIS1 remained in the same position relative to 1/HindIII linear DNA under different switching conditions (Fig. 2, 3 and 4), which clearly shows that it moves as linear DNA (Fig. 4). Although the wells in Fig. 4 were loaded with equal plug sizes, the band corresponding to the supercoiled form decreased in intensity and the brightness of the well area increased with progressive exposure to UV light (lanes 8 and 9), this suggests that the DNA remaining in the well is circular and cannot enter the gel matrix. Narayanan et al., (1993) studied the migration of supercoiled and relaxed rice mitochondrial DNA in CHEF gels and confirmed that relaxed circular DNA molecules failed to enter pulsed-field gels. Levene & Zimm (1987) suggested that relaxed circular DNA is ensnared by the gel fibers, especially at high field strengths. In this study, no CCC plasmid DNA was revealed from the different aquatic Streptomyces isolates using conventional gel electrophoresis. However, gentle lysis of Streptomyces cells in LMP agarose and subjecting of DNA to PFGE (CHEF-DR II) revealed high molecular-weight plasmids. REFERENCES Bentley, R. & R. Meganathan (1981). Geosmin and methylisoborneol biosynthesis in streptomycetes. FEBS Letters, 125:220-222 Chardon-Loriax, I., M. Charpentier & F. Percherson (1986). Isolation and characterization of a linear plasmid from Streptomyces rimosus. FEMS Microbiol. Lett., 35:151-155 Gerber, N.N. (1979). Volatile substances from actinomycetes: their role in odor pollution of water. Crit. Rev. Microbiol., 7: 191- 214 Katz, E., C.J. Thompson & D.A. Hopwood (1983). Cloning and expression of the tyrosinase gene from Streptomyces antibioticus in Streptomyces lividans. J. Gen. Microbiol., 129:2703-2714 Hayakawa, T., T. Kanaka, N. Sakaguchi, N. Otake & H. Yonchara (1979). A linear plasmid-like DNA in Streptomyces sp. producing lankacidin group antibiotics. J. Gen. Appl. Microbiol., 25:255-260
Hirochika, H., K. Nakamura & K. Sakaguchi (1982). Analysis of linear pla- smids isolated from Streptomyces: association of protein with the ends of the plasmid DNA. Plasmid, 7:59-65 Hirochika, H., K. Nakamura & K. Sakaguchi (1984). A linear DNA plasmid from Streptomyces rochei with an inverted terminal repetition of 614 base pairs. EMBO J., 3:761-766 Hopwood, D.A., M.J. Bibb, K.F. Chater, T. Kieser, C.J. Burton, H.M. Kieser, D.J. Lydrate, C.P. Smith, J.M. Ward & H. Schrempf (1985). Genetic Manipulation of Streptomyces. A Laboratory Manual. The John Innes Foundation, Norwich Ishibashi, Y. (1992). Genetic studies into musty odor production by acti- nomycetes. Water Sci. Tech., 25:171-176 Izaguirre, G., C.J. Huang, S.W. Krasner & E.G. Means (1982). Geosmin and 2-methylisoborneol from cyanobacteria in three water supply systems. Appl. Environ. Microbiol., 43:708-716 Keen, C.L., S. Mendeloritz, G. Cohen, Y. Aharonowitz & K.L. Roy (1988). Isolation and characterization of a linear DNA plasmid from Streptomyces clavuligerus. Mol. Gen. Genet., 212:172-176 Kieser, T. (1984). Factors affecting the isolation of ccc DNA from Streptomyces lividans and Escherichia coli. Plasmid, 12:19-36 Kinashi, H. & M. Shimaji (1987). Detection of giant linear plasmid in antibiotic producing strains of Streptomyces by the OFAGE technique. J. Antibiotics, 40:913-916 Leven, S.D. & B.M. Zimm (1987). Separation of open circular DNA using pulsed-field electrophoresis. Proc. Natl. Acad. Sci. USA, 84:4054-4057 Marcina, F.L., D.J. Kopecko, K.R. Jones, D.J. Ayers & S.M. McCowen (1978). A multiple plasmid-containing Escherichia coli strain: Convenient source of size reference plasmid molecules. Plasmid, 1:417-420 Narayanan, K.K., C.P. Andre, J. Yang & V. Walbot (1993). Organization of a 117-kb circular mitochondrial chromosome in IR36 rice. Curr. Genet., 23: 248245 Okanishi, M., T. Ohta & H. Umezewa (1970). Possible control of formation of aerial mycelium and antibiotic production in Streptomyces by episomic factors. J. Antibiotics, 23:45-47. Redshaw, P.A., P.A. Mccann, L. Sankaran & B.M. Pogell (1976). Control of differentiation in streptomycetes: Involvement of extrachromosomal deoxyribonucleic acid and glucose repression in aerial mycelium development. J. Bacteriol., 125:698-705 Redshaw, P.A., P.A. Mccann, M.A. Pentella & B.M. Pogell (1979). Simultaneous loss of multiple differentiation in aerial mycelium-negative isolates of Streptomyces. J. Bacteriol., 137:891-899 Saadoun, I.M., K.K. Schrader & W.T. Blevins (1993). Genetic determinants of earthy off-flavor production by aquatic species of Streptomyces and Lyngbya. Gen. Meet. Am. Soc. Microbiol., Abstr. N-69. p. 309 Schrader, K.K. & W.T. Blevins (1993). Geosmin-producing species of Streptomyces and Lyngbya from aquaculture ponds. Can. J. Microbiol., 39, 834-840 Sermonti, G., L. Lanfaloni & M.R. Micheli (1980). A jumping gene in Streptomyces coelicolor A3(2). Mol. Gen. Genet., 177:453-458 Tabachek, J.L. & M. Yorkowski (1976). Isolation and identification of blue- green algae producing mud odor metabolites, geosmin, and 2-methylisoborneol, in saline lakes in Manitoba. J. Fish. Res. Bd. Canada, 33:25-35 Copyright 1997 C.E.T.A., The International Centre for Theoretical and Applied Ecology, Gorizia The following images related to this document are available:Photo images[ac97010c.jpg] [ac97010b.jpg] [ac97010a.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}