 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Annals of African Medicine, Vol. 8, No. 1, March, 2009, pp. 1-9 Transmission Dynamics Of Malaria In Four Selected Ecological Zones Of Nigeria In The Rainy Season 1,2 O. O.Okwa, 1F.I. Akinmolayan, 2V.Carter and 2H.Hurd 1Department of Zoology, Lagos State University, Apapa, Lagos, Nigeria Accepted: 17 August 2008 Code Number: am09001 Abstract Background: Two of the problems of malaria parasite vector
control in Nigeria are the diversity of Anopheline vectors and large size of
the country. Anopheline distribution and transmission dynamics of malaria were
therefore compared between four ecotypes in Nigeria during the rainy season. Key words: Malaria; vectors; Plasmodium falciparum; transmission dynamics; ecotypes Résumé Fond: Deux des problèmes de la commande de vecteur de parasite de
malaria au Nigéria sont la diversité des vecteurs d'Anopheline et grande taille
du pays. La distribution d'Anopheline et la dynamique de transmission de la
malaria ont été donc comparées entre quatre ecotypes au Nigéria pendant la
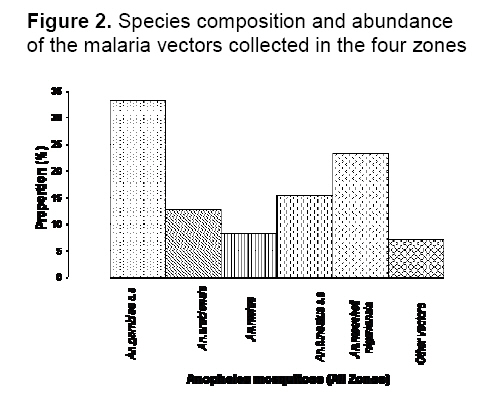
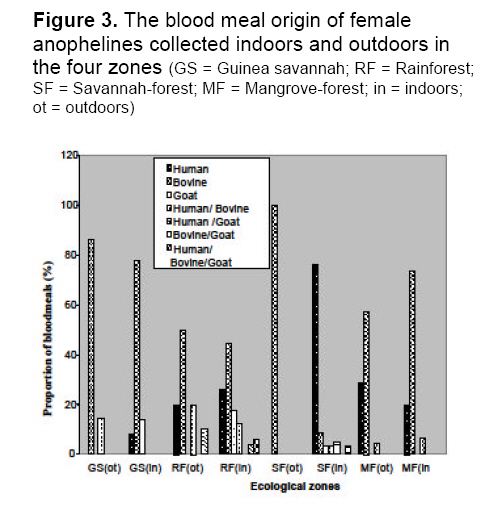
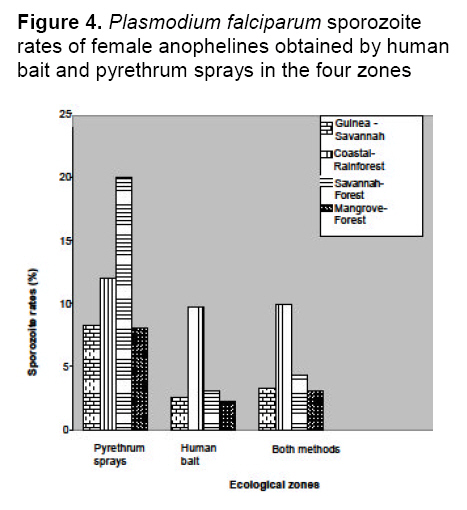
saison des pluies. Mots clés: Malaria, vecteurs, Falciparum de Plasmodium, dynamique de transmission, ecotypes Malaria is holoendemic in Nigeria, accounting for 25% of infant mortality and 30% of childhood mortality.1 Ninety five percent of malaria infections in Nigeria are caused by Plasmodium falciparum and five percent by Plasmodium malariae. 2 According to Gallup and Sachs, 2 malaria transmission is however geographically specific. De Mellion3 also reported that malaria vectors exhibit behavior variations in different localities. The knowledge of major vectors and their bionomics in Africa remains a problem.4 As a focal disease, malaria will therefore differ in its characteristics from place to place, since all malaria vectors do not exhibit identical behavior and ability to transmit parasites. Hay et al5 reviewed entomological inoculation rates (EIR) across Africa but there was no data from Nigeria. Nigeria is a large and diverse country and transmission dynamics will vary according to climatic, geographic and socio-economic conditions. Large areas of the country have no reliable data on the presence or absence of vectors and there is little information on sporozoite rates in southern Nigeria.6,7 Most of the original information on the transmission of malaria in Nigeria comes from isolated and intermittent studies in the northern parts of the country,8 In southern Nigeria, a few localized and short-term studies of mosquito populations were made9 and more recently Awolola et al 7, 8 studied population dynamics and biting behavior of anophelines in this area. Onyabe and Conn10 reported the distribution of two major malaria vectors, An. gambiae s.s and An. arabienisis, in southwestern Nigeria and most recently Awolola et al11 focused on the An. funestus group. In Nigeria, large numbers of Anopheles species are thought to be involved in transmission, but their distribution and vectoral capacity is not fully understood. Species complexes that vary in behavior and vectorial capacity present a real problem to malaria control.12 All the vectors belong to species complexes whose members vary widely in their vectorial capacity and competence.13 Correct analysis of the distribution of specific malaria vectors is one of the prerequisites for meaningful epidemiological studies and for planning and monitoring of successful malaria control or eradication programmes.14 A regular assessment of each country's malaria situation is worthwhile because control measures can only be effective if the abundance, behavior and proportion of the species are known. It is therefore very important to understand the dynamics of the transmission of malaria in a large country like Nigeria with different ecological zones. The aim of this study was to provide information on, and compare, species distribution, blood meal sources, sporozoite rates and entomological inoculation rates (EIR) of vectors in four ecological zones of Nigeria. This knowledge will facilitate better understanding of the dynamics of malaria transmission and could lead to development of early warning systems and species-specific vector control activities. Materials and Method Study areas Nigeria is approximately 923,768 sq. km (Figure 1). There are seven ecological zones in Nigeria, the arid savannah in the north gradually turning to humid forest in the south.10 Rainfall is the real climatic variable in Nigeria with June to September the rainiest months throughout the country. Mosquitoes were collected from the following four ecotypes based on accessibility, logistics, personnel and ethical reasons. The study sites were selected randomly based on the cooperation of people. Northwestern focus (Northern Guinea Savannah zone) Bungudu-Gusau, Zamfara State, where the Hausa is the predominant ethnic group, falls within the Northern Guinea Savannah zone of northern Nigeria. The rural population is mainly agricultural. Rainfall is approximately 500-700 mm per year. The numerous burrow pits; cattle dips, quarries, communal laundries and water storage containers provide small standing water sources that contribute to perennial breeding sites. Cattle are mainly found in compounds together with goats, sheep, dogs, fowl and bats Southwestern focus (Rainforest zone) Badagry, Lagos State, is a coastal suburban town, close to the Atlantic Ocean. The predominant ethnic groups are the Eguns and Yorubas. Fishing, poultry farming and trading are the basic occupation. The area is characterized by swampy sandy soil, which becomes waterlogged during the rainy season. Rainfall is approximately 2000- 3000 mm per year. Goats, sheep, pigs, fowls and dogs were common animals. Cattle are also found in the abattoirs and ranches. Southeastern focus (Savannah - forest zone) Onitsha, Anambra State, is in a sub-urban inland region. It falls within the savannah - forest zone in east central Nigeria. The predominant ethnic group is the Igbos. They are mainly involved in trading. The area is overpopulated because of the commercial activities. Rainfall is approximately 1900 mm per year. Dogs, fowl, goats, sheep, rabbits and cats are the common animals. Cattle were found at the abattoirs and cattle ranches. South-southern focus (Mangrove forest zone) Bonny, Rivers State is also a coastal suburban town that falls within the mangrove rainforest zone. It is within the Niger delta area very close to the Atlantic Ocean. The predominant ethnic groups are the Ijaws, who are mainly fishermen. Rainfall is approximately 2000- 4000 mm per year. Cats, fowl, dogs and goats are common animals. Cattle were found at the abattoirs and cattle ranches. Mosquito collection Collections were made during the rainy season (once a month) between July and October 2005 in the four zones. Female mosquitoes were caught outdoors between 1800-0600 hours by two human bait collectors who worked alternatively. Outdoor collections were made around open gutters, pools of water, open containers and abandoned tires and vehicles. They interchanged after 6 hours. An aspirator was use to collect the mosquitoes that landed and attempted to feed. Mosquitoes were also caught by pyrethrum spray indoors in the evenings between 1800- 2000 hours by floor sheet collections. The Mosquitoes were preserved in 70% ethanol. Morphological identification of mosquitoes Anopheline mosquitoes were distinguished from Culicines according to the morphological characteristics of their maxillary palps using the morphological keys of Gilles and Coetzee.15 The distinguishing feature of An. m. nigeriensis being a fringe spot opposite vein 6 on the wing was used to distinguish it from other anophelines.16 Dissection of mosquitoes All the female anophelines collected were dissected. They were cut transversely at the thoraxes between the 1st and 3rd pairs of legs under a dissecting microscope (x20). The abdomens of blood-fed anophelines were used for blood meal analysis while the heads and thoraxes were used for sporozoite detection. The wings and legs were used for species identification following DNA extraction. Molecular identification of Anophelines DNA Extraction Genomic DNA from whole male mosquitoes and wings and legs of female mosquitoes were extracted according to the standard procedures of Collins et al17 Extracted DNA was resuspended in 50 ml of PCR grade water. Polymerase chain reaction PCR was performed with universal and species specific primers for the An. gambiae and An. funestus species complexes. Molecular identification of An. gambiae species complex is based on the species-specific nucleotide sequences in the ribosomal DNA intergenic spacers (IGS) as described by Scott et al18 The sequences of the An. gambiae complex primers used were as follows: Universal 5¢-GTGTGCCCCTTCCTCGATGT-3¢, An. gambiae 5¢-CTGGTTTGGTCGGCACGTTT-3¢, An. melas 5¢-TGACCAACCCACTCCCTTGA-3¢ and An. arabiensis 5¢-AAGTGTCCTTCTCCATCCTA-3¢. PCR for An. gambiae complex consisted of one cycle of initial denaturation at 94°C for 5 minutes, 30 cycles of denaturation at 95°C for 30 seconds, annealing at 50°C for 30 seconds, extension at 72°C for 30 seconds and final extension at 72°C for 7 minutes. For the An. funestus species complex, molecular identification was based on species–specific primers in the internal transcribed spacer region (ITS2) on the ribosomal DNA as described by Koekemoer et al19 The sequences of the An. funestus complex primers used were: Universal 5¢-TGTGAACTGCAGGACACAT-3¢, An.funestus 5¢-GCATCGATGGGTTAATCATG-3¢, An. vaneedi 5¢-TGTCGACTTGGTAGCCGAAC-3¢, An.rivulorum 5¢-CAAGCCGTTCGACCCTGATT-3¢, An.parensis 5¢-TGCGGTCCCAAGCTAGGTTC-3¢ and An. leesoni 5¢-TACACGGGCGCCATGTAGTT-3¢. PCR performed for An. funestus complex consisted of one cycle of initial denaturation at 94 °C for 2 minutes, followed by 30 cycles of denaturation at 94°C for 30 seconds, annealing at 45°C for 30 seconds and extension at 72°C for 40 seconds, with additional final extension at 72°C for 5 minutes. Laboratory strains of An. gambiae and An. funestus were used as controls. Amplification was performed in a Gene AMP PCR system 9700 using Sigma (U.K) reagents throughout. Oligonucleotides were produced by Proligo (France). All PCR products were electrophoresed on a 1.4% ethidium bromide agarose gel. The amplified fragments were then visualized by UV transilluminator and documented using Syngene bio-imaging system. Identification of blood meal origin A direct ELISA using anti-phosphatase conjugates; anti-human IgG (Fab specific); anti-bovine IgG (whole molecule) and anti-goat IgG (whole molecule) [Sigma], were used to identify human, cattle (bovine) and goat (ovine) host blood respectively, based on the procedures of Beier et al20 A total of 4160 mosquitoes with blood meals were assayed for each host in 96-well flat-bottomed well plates with absorbance read 30 minutes after the addition of substrate (pNPP) at 405nm on a Labsytem Multikans Multisoft Type 349 ELISA reader. Samples were considered positive if the optical densities (OD) were at least twice the mean of four negative wells on the same plate.21 Positive controls were female mosquitoes with known blood meals. Negative control contained male mosquito triturates. Human blood index The human blood index (HBI), which is the proportion of female anophelines giving a positive reaction for human blood alone or multiple blood feeds with human blood, was calculated.22 This is the percentage of female anophelines that had human blood or mixed blood that contained human blood, i.e., No. of female anophelines with human blood or mixed human blood/No. of female anophelines tested Plasmodium falciparum sporozoite rate The heads and thoraxes of individual mosquitoes were tested for P. falciparum circumsporozoite protein (CSP) using Pf2A10 monoclonal antibody, as recommended and modified by Wirtz et al.23 The monoclonal and peroxidase-conjugated antibodies were obtained from Centre for Disease Control (CDC, Atlanta, Georgia). A sandwich ELISA was carried out on dried mosquitoes. The heads and thoraxes were ground individually in 50 ml grinding buffer containing, bovine serum albumin (2.5g), boiled casein (1.25g) in phosphate buffered saline (250mls), phenol red (100ul) and Nonidet P-40 (1,250ul) and the final volumes of each triturate brought to 250 ml. These triturates (50 ml) were then used in the ELISA test according to the standard protocols of Beier et al.20 Five negative controls (male mosquitoes) and three positive controls (the appropriate synthetic peptide) were included in each 96-well flat-bottomed microtiter plate. The OD was read at 415nm, 60 minutes after the addition of substrate (ABTS). Any sample giving an OD that was at least twice that of the mean OD for the negative controls on the same plate was considered CSP positive.21 Positive samples were retested for confirmation. Sporozoite and entomological inoculation rates Sporozoite rates were determined as the percentage of mosquitoes carrying P. falciparum CSP antigen. Combined data from mosquitoes collected by human bait and pyrethrum spray during the whole study were used to calculate sporozoite rates of each species in each area. However, only data from the human bait collections were used to calculate the entomological inoculation rates (EIR) as the product of the sporozoite and the man-biting rate; the man-biting rate being the number of mosquitoes biting per person per night .i.e. per human bait. EIR is expressed as infective bites per person for the rainy season (ib/p). Statistical analysis χ2 was used to determine the differences in results between the study sites and among species. P<0.05 was regarded as an acceptable level of significance. Results Species distribution A total of 16, 410 anophelines were collected from the four foci. Pyrethrum sprays collected more mosquitoes. Five species were identified; three were from the An. gambiae complex but of the An. funestus complex, only An. funestus s.s was present. Overall, An. gambiae s.s was the most abundant species and this was followed by An. moucheti nigeriensis. The least common species identified was An. melas, which was only found in two zones. In addition, 7% of the collection was un-identified (Figure 2, Table 1). In the Guinea savannah, out of 2870 anophelines, only three species were identified. There were more An. funestus s.s than An. gambiae s.s or An. arabiensis and no An. m. nigeriensis or An. melas were found, but there were other secondary species. In the coastal rainforest, all five species were identified in a collection of 5580 anophelines. An. gambiae s.s and An. m. nigeriensis were the most abundant; both 31.8% of the collection. An. funestus s.s, An. arabiensis, and An. melas, were also identified and 7.31% were unidentified species. In the savannah-forest, out of 4050 anophelines, four species were identified; An. melas being absent, whereas, in the mangrove–forest four species were also identified but An. funestus s.s. was not found and 5.82% were unidentified. See Table 1 for values. Species distribution between zones differs significantly X2=487.8 df=5 P<.05. Biting females Of the 1110 biting females collected by human bait outdoors, An. gambiae s.s predominated at 37% of the total catch; An. m. nigeriensis represented 28%, An. funestus s.s 15.3%, An. arabiensis 11% An. melas 3.6%. In terms of distribution, the highest numbers of biting species were found in the Guinea savannah where biting An. gambiae s.s were 41.7% of the catch for that area. This was followed by the rainforest where biting An. moucheti nigeriensis were 24.6% of the catch, savannah-forest with An. gambiae s.s at 40% and Mangrove–forest where biting An. moucheti nigeriensis were 48.6% of the catch (Figure 3). Blood meals A total of 4160 (25.3%) anophelines were blood fed. Overall, 1360 (32.7%) had fed on bovids, 820 (19.7%) on humans and 300 (7.21%) had mixed feeds with human blood as a component. Only 150(3.6%) goat and 40(0.96%) bovine/goat meals were identified (overall human blood index of 27%) (Table 2). Generally, more blood meals had been taken from bovid. However, in the savannah forest 440 (73.3%) of blood meals were taken on humans alone. The highest proportion of bovid blood meals was in the Guinea savannah (79.6%) of the 1230 blood fed females examined. In this zone, 140 (58.3%) of outdoor caught-females contained blood meals and of these 120 (86%) contained blood meals of bovid origin and 20 (14.3%) mixed bovid and human meals. Indoors in the Guinea savannah, 390 (78%) blood meals were of bovid origin and 40 (8%) of human origin. There were no multiple blood feeds (Figure 4). A human blood index of 4.87% was recorded here (Table 2). In the rainforest, 40% of the 1240 blood fed females was caught outdoors by human bait catches and 50% of these had blood meals of bovid origin only, whereas only 20% were of human origin, 20% human/bovid origin and 10% from bovid/goat origin. A similar picture emerged for indoor pyrethrum catches in this zone (Figure 4). A human blood index of 32.2% was obtained (Table 2). In the Savannah-forest, from 890 blood fed females, few females were found outdoors and they were all bovine fed, whereas 76% blood meals from indoors-caught anophelines were of human origin and only 50 (8.6%) of the blood meals were taken on bovid. There were few multiple blood feeds of human/bovid 20 (3.45%), human/goat 30 (5.2%) and human/bovid/goat 20 (3.45%). Human blood index of 57.3% was recorded in the rain forest (Table 2). In the mangrove- forest, there were 800 blood fed females. Seventy (19%) were caught by human bait outdoors, out of which 20 (28.5%) had taken blood meals from a human, 10(4.2%) from human/bovid and 40 (57.1%) from bovids. indoors, 330 (73.3%) of the blood meals were taken from bovids and 90 (20%) on human. A human blood index (HBI) of 18.75% was recorded (Table 2). HBI between zones differs significantly (p <.05). Sporozoite rates Out of a total of 11 990 female anophelines examined for Plasmodium falciparium sporozoites, 670 (5.6%) were positive and all species identified contained sporozoites and thus acted as malaria parasite vectors in at least one zone (Table 3). A higher proportion of anophelines caught by the pyrethrum spray method contained sporozoites than by human bait. In the Guinea savannah, 50 (2.6%) female anophelines caught indoors and 20 (8.3%) caught outdoors had sporozoites. This gave an overall sporozoite rate of 3.24% out of the 2160 females examined. An. funestus had the highest sporozoite rate of 2.3% in this zone. In the rainforest, a sporozoite rate of 12% outdoors and 9.71% indoors was recorded for the female anophelines examined. This indicated an overall sporozoite rate of 9.9%, which was the highest among the four zones with An. gambiae, s.s having the highest sporozoite rate of the five species identified (4.3%). In the savannah- forest, a sporozoite rate of 20% outdoors and 3.1% indoors was recorded from 3510 female anopheles examined. This gave a sporozoite rate of 4.3%, with An. gambiae s.s again having the highest sporozoite rate of 2.3%. In the mangrove-forest, there was a sporozoite rate of 8.11% outdoors and 2.3% indoors. This gave an overall sporozoite rate of 3.11%, which was the lowest obtained among the zones. An. gambiae s.s and An. moucheti nigeriensis had the highest sporozoite rates of 1.2% each for this zone. (Table 3). Sporozoite rates between species differs significantly (p<.05). Sporozoite rates between zones differs significantly (p<.05). Entomological inoculation rates (EIR) Overall for the whole of Nigeria, mean EIR calculated from human bait catches alone was 13.6 ib/p for the rainy season. The EIR was highest in the southern rainforest (24.7 ib/p) and lowest in the drier northern Guinea savannah with 7.7ib/p (Table 4). Table 1. Species composition and abundance of anophelines collected in the four ecological zones of Nigeria in the rainy season
a Species distribution between zones differs significantly; X2=487.8 ; df=5; p<.05; GS = Guinea Savannah; RF= Rainforest; SF = Savannah-forest; MF = Mangrove- forest Table 2. Host preferences of blood fed malaria vectors and their human blood index in four zones
a Human blood index (HBI) between zones differs significantly; X2 =47.26; df =3; p<.05; HBI is the proportion of anophelines with human blood and multiple blood feeds with human out of the total blood meals taken Table 3. Proportion of female anophelines with sporozoites in their salivary glands in the four zones
a Sporozoite rates between species differs significantly;
X2 = 41.9; df = 5; P <.05 Table 4. Entomological inoculation rates (EIR) of female anophelines obtained by human bait (HB) in the four zones in the rainy season July-October, 2005
ainfective bite per person per rainy season; GS = Guinea savannah; RF = Rain forest; SF =Savannah forest; Discussion This study investigated the principal vectors of the malaria parasite in Nigeria and their role in malaria transmission. Species distribution in the four areas suggests that members of An. gambiae, An. moucheti nigeriensis and An. funestus complexes can be found in sympathy. Several authors15, 16 also reported this sympatric distribution. In this study, An. gambiae s.s was distributed over the four zones, and was most abundant, which is consistent with findings of Bruce-Chwatt24 who concluded that An. gambiae s.s is omnipresent in Nigeria, because of its indiscriminate breeding habitats. They described it as highly endophilic, anthropophagous, wet season vector, but can occasionally be zoophilic and exophilic. Although we observed that An. gambiae s.s fed on non-human hosts, it had the highest P. falciparum sporozoite rate of 2.33%. An. arabiensis has been described as a savannah vector, in isolated populations, deforested areas and predominant in the dry season.8 In this study, An. arabiensis was found in considerable numbers in the rainforest zone. Wherever An. arabiensis occur in the rainforest, it is associated with a history of extensive land clearance.12 The coastal sampling sites in this study were areas where deforestation is increasing due to urbanisation. Onyabe and Conn10 reported that there has been an extension in the range of An. arabiensis in Nigeria, prevailing in arid zones, but also in some forest zones. Puzzling shifts in species composition of An. arabiensis and An. gambiae s.s have been observed in Nigeria.10 Githeko et al25 reported that An. arabienisis could be anthropophagous, where there are less animal hosts, as we observed, in the Savannah- forest, where An. arabiensis was responsible for 34.1% of human blood meals. An. arabiensis appears to be a good vector of malaria, especially in the Savannah-forest. An. melas occurred only in the Coastal rainforest (7.65%) and Mangrove forest (24.6%). According to Bruce-Chwatt, 24 this is the dominant vector in the west coast being related more to sea tides. De Mellion3 first reported it from Lagos. Until recently, An. funestus s.s, has received scant attention.11 This is inconsistent with its obvious major role in malaria transmission, with a sporozoite rate of 1.42%, second only to An. gambiae s.s. Most research has focused on the members of An. gambiae complex. According to Gilles and Cozetee, 16 the An. funestus group may be as complex and problematic as the An. gambiae group with different biology and vectorial capacity. In some areas of Nigeria, it has been projected that it could replace An. gambiae s.s as the major vector of endemic malaria. Three members of this group have been found to date in Nigeria: An. funestus s.s, An. leesoni and An. Rivulorum.7, 11 However, we only identified An. funestus s.s in three ecotypes. We did not observe An. funestus in the mangrove forest focus. This could be because of the more restricted habitat choice of An. funestus s.s preferring very clean fresh shaded water. This explains, in part, why malaria transmission was less intense in the mangrove forest than in the rainforest. An. moucheti nigeriensis was identified only in the Southern zones in this study, but was more abundant in the rainforest. An. moucheti as a species complex found in sympatry with An. gambiae complex and a major or only human malaria vector in villages and towns situated in forest areas. An. moucheti was reported as an important primary vector in Nigeria, 15 but played a lesser role when compared to An. gambiae s.s and An. funestus s.s.). This is consistent with the present study in which An. moucheti nigeriensis has a lower sporozoite rate of 0.75%. Despite this epidemiological importance, very few studies have been carried out on this vector. PCR identification should be extended to other important anopheline species complexes such as An. moucheti. Bruce-Chwatt24 reported 28 anopheline species in Nigeria and in the same year, De Mellion3 reported an increase in number of anophelines, infected in nature and mainly exophilic. Recently, Awolola et al6 stated that the diverse species of anophelines involved, confounds studies on malaria transmission in Nigeria. We obtained a sporozoite rate of 0.33% for secondary vectors, showing their role in transmission. In Northern Nigeria with the shortest transmission period, one expects to get epidemic malaria instead of holoendemic, but there is no evidence of such to date. The presence of cattle within compounds must have attracted An. arabiensis to the vicinity of humans. This was reflected by low a HBI and EIR in the Guinea savannah. In a coastal region of southwestern Nigeria, Awolola et al6 reported the presence of the following vectors: An. gambiae s.s, An. moucheti, An. melas and An. arabiensis. We also identified all of these vectors in the southwestern focus in addition to An. funestus s.s. In this study, An. moucheti is an important vector in the mangrove with a sporozoite rate of 8.1% outdoors. The highest HBI (57. 3%) was recorded in the savannah- forest zone, which was overpopulated with humans. On several occasions, we detected multiple blood meal sources. These multiple blood meals suggest that female anophelines take successive bites to complete a blood meal. To complete their gonotrophic cycle, female anopheline mosquitoes usually require a second blood meal one day after the first to mature the first egg batch.15 Shililu et al22 and Githeko et al,25 in two different studies in Kenya, also reported that anophelines took multiple blood feeds of human, bovid and avian origin. In their study, some blood meals proved negative for all tested hosts, as was also found in this study. According to Bruce- Chwatt, 24 the mean overall sporozoite rate in Nigeria is 6%. This is close to the estimated sporozoite rate of 5.6% that we obtained. We obtained overall sporozoite rates for An. gambiae s.s. (2.33%) and An. funestus s.s. (1.42%). Beier 27 also reported that sporozoite rate of An. gambiae s.s and An. funestus s.s in tropical Africa is about 1%-5%. The mean EIR indicates that in Nigeria, about 13 infective Anopheles could transmit malaria parasite successfully in the rainy season (July –October). The malaria problem in Africa south of the Sahara represents a peculiar case because the vectorial system is the most complex anywhere. Beier 27 also suggested that malaria transmission dynamics is variable throughout Africa with huge variability in transmission patterns even within villages few kilometres apart. This vectorial system diversity will impact on malaria epidemiology and control. The An. gambiae complex is not the only vector in the field. Targeting only this species by whatever method is nonsense. This study expands the view that the malaria vectorial system in Nigeria is more complex than expected, looking at the combined contribution of these mosquito species to malaria transmission. The diversity of the epidemiological situation within the country ecotypes presents differing malaria situation. Comprehensive knowledge of behavior and heterogeneities that exist within, and among these vectors, will benefit the whole country. Any strategy aiming at control will have to account for this heterogeneity. Acknowledgments This work was supported by a Commonwealth Fellowship grant to Dr Omolade Okwa tenable at Keele University, UK. Centre for Disease Control, Atlanta, Georgia, U.S.A is acknowledged for the provision of the antibodies. We thank Dr Lynn Mc Carroll of the Liverpool School of Tropical Medicine for her assistance with the polymerase chain reaction. Prof Chris Curtis and Ms Shahida Begum of the London School of Hygiene and Tropical medicine are acknowledged for their assistance with the sporozoite ELISA. References
Copyright 2009 - Annals of African Medicine The following images related to this document are available:Photo images[am09001f1.jpg] [am09001f3.jpg] [am09001f2.jpg] [am09001f4.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}