 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Acta Botanica Sinica, Vol. 45, No. 7, 2003, pp. 762-765 Rapid Communication Identification of Oryza Species with the CD Genome Based on RFLP Analysis of Nuclear Ribosomal ITS Sequences1* BAO Ying1, 2, GE Song (1. Laboratory of Systematic and Evolutionary
Botany, Institute of Botany, The Chinese Academy of Sciences, Beijing
100093, China; 2. Department of Biology, Qufu Normal University, Qufu
213765, China)
Supported by the National Natural Science Foundation of China (30025005) and The Chinese Academy of Sciences (KSCXZSW-101A). Received: 2002-11-07 Accepted: 2003-03-17 Code Number: as03002 ABSTRACT: The genus Oryza consists of approximately 24 species with 10 recognizedgenome types (A, B, C, BC, CD, E, F, G, HJ, HK). The species with the CD genome are endemic to the Central and South America and comprise three species, i.e. O. latifolia Desv., O. alta Swallen and O. grandiglumis (Doell) Prod. Although the three species can be easily distinguished from the species with the other genomes in Oryza by their morphological characteristics, it is very difficult to distinguish them from one another. Recent studies suggested that O. alta and O. grandiglumis should be the same species (O. grandiglumis), whereas O. latifolia retains its taxonomic position unchangeably. In this paper, a total of 77 clones of the nuclear ribosomal internal transcribed spacer (ITS) from 11 samples representing different geographical races were sequenced. Using DNA Strider 1.2 software, the restriction enzyme digestion sites of these clone sequences were analyzed. Based on restriction fragment length polymorphism (RFLP) of ITS sequences, a method toidentify the species with CD genomein Oryza was proposed. The method is rapid and convenient and all experimental procedure includes only three steps: (1) to amplify the ITS fragment with the routine primers; (2) to digest PCR products with restriction enzymes FokI and Dra III or both; (3) to run the digested product on 1% agarose gel and identify the sample based on the restriction profiles. Key words: Oryza ; identification; CD genome; RFLP; ITS The genus Oryza consists of approximately 24 species with 10 recognized genome types (A, B, C, BC, CD, E, F, G, HJ, HK)(Ge et al, 1999). The species with the CD genome are endemic to the Central and South America and comprise three species, i.e. O. latifolia, O. alta and O. grandiglumis (Vaughan, 1989). These species are perennial with erect and higher plants as well as big and open panicles (Nayar, 1973). Of them, O. latifolia is widely distributed in Central and South America and on the Caribbean islands. In contrast, O. alta and O. grandiglumis occur only in South America (Vaughan, 1989). In the genus Oryza, the species with the CD genome are easily distinguished from the species with other genome types by their larger plants, broader leaves (more than 2 cm) and unique geographical distribution (Vaughan, 1989). However, it is difficult to distinguish these three species from one another. Up to date, different opinions have been raised regarding whether the three species are treated as separate or a single species (Nayar, 1973; Jena and Kochert, 1991; Buso et al, 2001). Many molecular studies using the nuclear and chloroplast gene sequences (Ge et al, 1999), AFLP markers (Aggarwal et al, 1999) and RFLP of repetitive sequences (Federici et al, 2002) all indicated that O. alta and O. grandiglumis were closer to each other than either to O. latifolia. Recently, we studied the CD genome species based on the sequences from two nuclear genes (Adh1 and Adh2), two choloplast genes (matK and trnL-trnF) and the nuclear ribosomal internal transcribed spacer (ITS), and found that the accessions sampled from O. alta and O. grandiglumis were always clustered together and separated clearly from O. latifolia on multiple-gene trees (unpublished). Therefore, we suggest that O. alta and O. grandiglumis should be treated as same species (O. grandiglumis), while O. latifolia remains a separate species. A correct classification is the premise for effective utilization of plant resources and germplasm, especially for wild rice which contains an enormous gene pool for the genetic improvement of rice cultivars (Vaughan, 1989; Ge et al, 1999). Here we present a rapid and convenient method to distinguish the species with the CD genome based on the RFLP profile of the nuclear ribosomal internal transcribed spacer (ITS). This method can facilitate identification of Oryza species with the CD genome, and can be a potential way for utilization in the wild rice germplasm collection. 1 MATERIALS AND METHODS
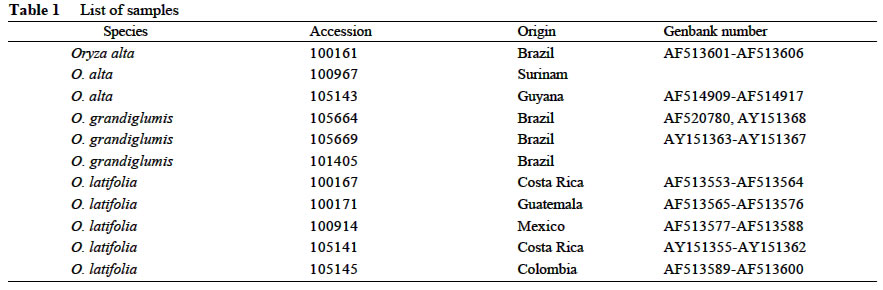
A total of 11 accessions were sampled in the study, including three accessions from each of Oryza alta Swellen and O. grandiglumis (Doell) Prod., five from O. latifolia Desv. The species names, accession numbers and origins are listed in Table 1. Plants were grown from seeds kindly provided by the International Rice Genebank at the International Rice Research Institute (IRRI) in the Philippines and developed in the greenhouse. Total DNA was isolated from fresh leaves of plants individually by using CTAB method (Doyle and Doyle, 1987). Nuclear DNA ITS region was amplified with primer ITS1 (5'-AGAAGTCGTAACAAGGTTTCCGTAGG-3') and ITS4 (5'-TCCTCCGCTTATTGATATGC-3') (White et al, 1990). Each amplification reaction (25 µL final volume) contains 10-20 ng template DNA, 10 mmol/L Tris-HCl (pH 8.3), 2.0 mmol/L MgCl2, 0.2 mmol/L of each dNTP, 5 pm each primer, and 0.75 units of Taq polymerase. In addition, 8% dimethylsulfoxide (DMSO) was added to the reaction mixture to facilitate denaturation during amplification (Baldwin et al, 1995). The PCR procedure includes 30 cycles of 1 min denaturation at 94 °C, 1 min annealing at 52 °C, and 1.5 min extension at 72°C and a final 10-min extension at 72 °C. Amplifications were performed in PTC-200 (PE) thermocycler. We have cloned and sequenced the internal transcribed spacer (ITS) for the CD genome species, including 14 clones for O. alta, seven clones for O. grandiglumis and 56 clones for O. latifolia. The ITS sequences of these species were deposited in the GenBank database under the accession numbers listed in Table 1. In order to obtain the diagnostic restriction profiles, we surveyed the restriction sites of the ITS sequences using DNA Strider 1.2 (Marck, 1988). The strategy for determining species-specific sites followed that of Ge et al (2001). For the purpose of confirmation of the diagnostic sites, 2 to 5 µL of PCR products of 11 accessions representing different geographical races were digested in each 10 µL reaction containing 1 U of restriction enzyme. Digested PCR products were electrophoresed on 1% TBE agarose gels. The gels were stained with ethidium bromide, visualized, and photographed under UV light. 2 RESULTS AND DISCUSSION
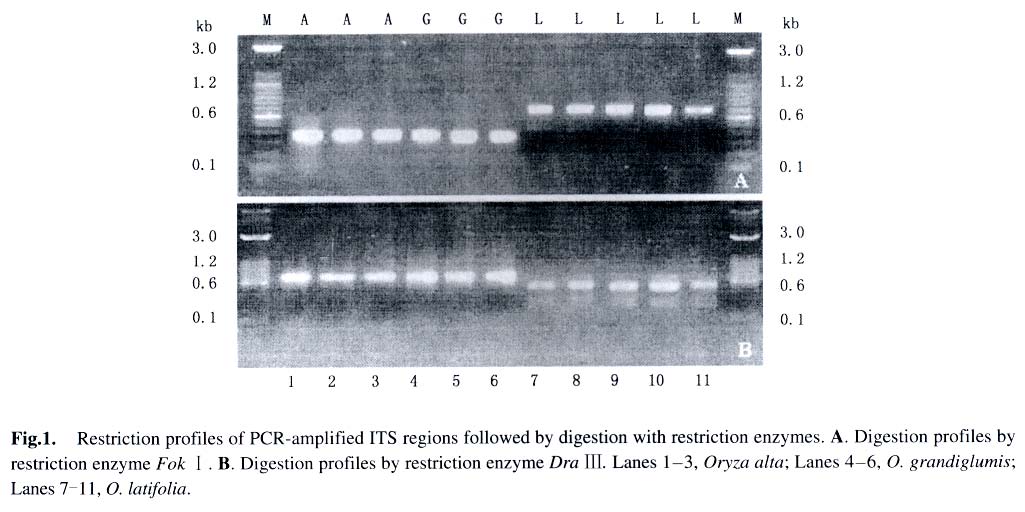
Using DNA Strider, we surveyed 170 restriction enzymes for the restriction sites of the ITS sequences of the CD genome species. Of them, the enzymes that had restriction sites conserved across accessions of the same species but variable among different species were chosen for the purpose of identifying species. So, two restriction enzymes of Fok Iand DraIII were chosen by which we could distinguish O. latifolia from O. grandiglumis (O. alta) unambiguously. However, no restriction site was found to distinguish O. grandiglumis from O. alta, demonstrating close relationship between them as indicated by many previous studies (Aggarwal et al, 1999; Ge et al, 1999; Federici et al, 2002). Restriction enzyme Fok I has no cutting site in the ITS region of O. latifolia but one cutting site in the middle of the ITS regions of O. alta and O. grandiglumis, resulting in a 700-bp band for O. latifolia and two nearly equal bands (~350 bp) for O. alta and O. grandiglumis. For restriction enzyme Dra III, on the contrary, one cutting site in O. latifolia but no cutting site in O. alta and O. grandiglumis were observed, resulting two bands (200 bp and 500 bp, respectively) for O. latifolia and one band (700 bp) for O. alta and O. grandiglumis. The restriction profiles of the two enzymes are shown in Fig.1. Consequently, we can distinguish O. latifolia from O. alta or O. grandiglumis clearly based on the restriction digestions of PCR-amplified ITS sequences by Dra III and Fok I. In summary, the method proposed in the present study is rapid and convenient. The experimental procedure includes only three steps: (1) to amplify the ITS fragment with the routine ITS primers; (2) to digest PCR products with restriction enzymes Fok I and Dra III or both; and (3) to run the digested product on 1% agarose gel and identify the sample based on the restriction profiles. The method can exclude the influence by artificial factors and improve validity of identification greatly. More importantly, this method offers an attempt to distinguish these species at any stage of their life cycle. REFERENCES:
Copyright 2003 - Acta Botanica Sinica. Free, full-text also available from http://www.chineseplantscience.com The following images related to this document are available:Photo images[as03002t1.jpg] [as03002f1.jpg] |
| |||||||||

{kind=link}
{kind=link}