 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
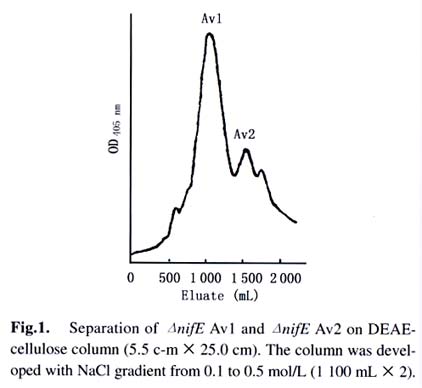
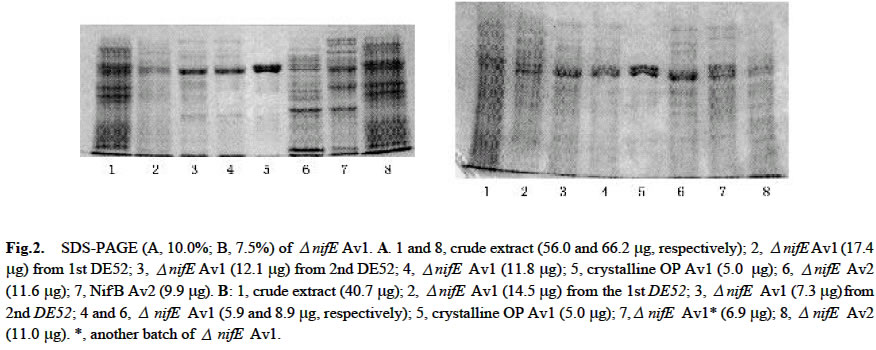
Acta Botanica Sinica, Vol. 45, No. 7, 2003, pp. 815-819 Purification and Activation In Vitro of MoFe Protein from a nifE Deleted Mutant Strain of Azotobacter vinelandii ZHAO Jian-Feng1, ZHAO Ying1, WANG Zhi-Ping2, LÜ Yu-Bing1, QIAN Zhong-Xing1, HUANG Ju-Fu1 * (1. Key Laboratory of Photosynthesis and Environmental Molecular Physiology, Institute of Botany,
The Chinese Academy of Sciences, Beijing 100093, China;
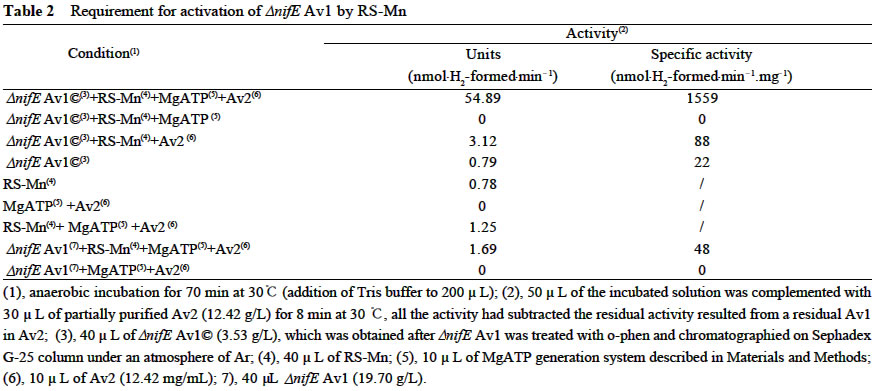
Received: 2002-06-18 Accepted: 2002-09-10 Code Number: as03010 ABSTRACT TheΔnif E MoFe protein (Δnif E Av1) was obtained by a chromatography on DE52, Sephacryl S-300 and Q-Sepharose columns from the unheated crude extract of nifE -deleted mutant strain (DJ35) of Azotobacter vinelandii Lipmann. The analysis by SDS-PAGE showed that the Δnif E Av1 was similar to OP MoFe protein (Av1) of A. vinelandii in the kinds and molecular weights of subunits (αand β subunit). When complemented with nitrogenase Fe protein (Av2), the Δnif E Av1 had hardly any proton-reduction activity, but could be significantly activated by FeMoco extracted from OP Av1. After theΔnif E Av1 was treated with an excess o-phenanthroline (o-phen) and chromatographied on Sephadex G-25 column under atmosphere of Ar, Δnif E Av1© was obtained. In the presence of both Av2 and MgATP regeneration system, the Δnif E Av1©, rather than Δnif E Av1, was significantly activated in vitro by a reconstituent solution containing Mn which composedof KMnO4, ferric homocitrate, Na2S, Na2S2O4(DT) and dithiothreitol (DTT). But in the absence of MgATP or Av2, the activation ofΔnifE Av1© did not happen. It indicates that activation ofΔnif E Av1 by RS-Mn requires the pretreatment with o-phen andthe simultaneous presence of Av2 and MgATP. Key words: Δnif E Av1; purification; activation and assembly in vitro ; FeMoco and reconstituent solution containing Mn The biological reduction of N2 is one of the most fundamental processes in nature. Mo-containing nitrogenase from Azotobacter vinelandii composed of MoFe protein (Av1) and Fe protein (Av2) plays a key role in the process. FeMoco in Av1 is thought to provide the site of substrate reduction. Biosynthesis of FeMoco involves the participation of six genes, nifQ, nifB, nifE , nifN, nifH and nifV (Tal et al, 1991). If one of the six genes is deleted or mutated, Av1 synthesized in vivo is one of Apo-Av1, FeMoco-deficient Av1. The Apo-Av1 can be activated in vitro by FeMoco extracted from Av1 (Shah and Brill, 1977; Pustian et al, 1989; Tal et al, 1991). The products of nifN and nifE genes function as a 200-kD α2β2 tetramer (NIFNE), which has been purified and partially characterized (Brigle et al, 1987). Although the exact role of NIFNE in FeMoco synthesis is unknown, it is thought to serve as a scaffold for the synthesis of Av1. It is shown that studies on mutant nitrogenases are of importance to further understand the mechanism of nitrogen fixation. To date, ΔnifH Av1 (from DJ54), NifBAv1 (from UW45) and ΔnifB Av1 (from DJ1143) have been purified and characterized, respectively, and the crystalline structure of the ΔnifB Av1(from DJ1143) was recently reported (Robinson et al, 1987; Puustian et al, 1989; Gavini et al, 1994; Schmid et al, 2002). In contrast with crude extract of Δ nif H strain, the maximum activities for the crude extracts of ΔnifB (from DJ141) and ΔnifE (from DJ35) strains activated by FeMoco are much less than that of wild-type (Tal et al, 1991). Recently, nif E- Rc1 was partially purified from a nifE - mutant of Rhodobacter capsulatus (Siemann et al, 2001). So, the purification, characterization and analysis of crystal structure of ΔnifE Av1 (from DJ35) are not only necessary, but also possible. Three genetically distinct nitrogenase systems exist in bacteria: the “classical” Mo-containing nitrogenase, a Vcontaining one and an “iron only” nitrogenase lacking both Mo and V. Like Mo-containing nitrogenase, the last two nitrogenases are composed of two separable proteins called component I (VFe protein and FeFe protein, respectively) and component II (Fe protein). A comparative study on the relationship between the structure and function of these MFe proteins (M is Mo, V and Fe, respectively) is of great importance for the elucidation of mechanism of nitrogen fixation (M üller et al, 1992). There are some preliminary evidence to postulate that n ew nitrogenases containing Mn or Cr could exist besides the well known three nitrogenases (Huang et al, 1994; 2001; 2002). However, nitrogenase MnFe protein and CrFe protein obtained by two different methods contained 20% -50% Mo. If FeMnco could be assembled in vitro and inserted into FeMoco-deficient ΔnifE Av1 using the previously described method (Huang et al, 1994), it is possible to obtain two new nitrogenases with lower ratio of residual Mo to Mn or Cr. This study was on the purification of ΔnifE Av1 and discovery of the possibility for its crystallization and its activation by a reconstituent solution containing Mn. 1 MATERIALS AND METHODS The nifE -deletion mutant strain of Azotobacter vinelandii Lipmann (DJ35) was kindly provided by Prof. Burgess. The construction and growth of DJ35 was reported by Brigle et al (1987). The cells of DJ35 were ruptured in French pressure cell at 1 000 psi. The solution of the ruptured cells was centrifuged for 30 min at 75 100g. The crude extract was applied to DE52 column (5.5 cm × 25.0 cm), followed by washing with 0.10 mol/L NaCl in an anaerobic Tris buffer and eluted with 0.50 mol/L NaCl in the buffer. The fraction from the column was diluted with 4-fold (V/V) buffer and applied to another DE52 column, then followed by developing with NaCl gradient from 0.10 to 0.50 mol/L. The separated ΔnifE Av1 and ΔnifE Av2 were diluted and applied to DE52 columns, respectively, then followed by eluting upstream with 0.50 NaCl in the buffer. The concentrated ΔnifE Av1 was further purified by chromatography on Sephacryl S300 column (2.5 cm × 72.0 cm) and Q-sepharose column (2.5 cm × 20.0 cm) which was developed with NaCl gradient from 0.10 to 0.30 mol/L, then followed by concentration on Q-sepharose column. The ΔnifE Av1 solution (134 mg) was adjusted to about pH 6.5, then followed by incubation with 32 mg of o-phen (256-fold over mole of protein) for 170 min at 30 °C under the anaerobic condition. The treated protein (ΔnifE Av1©) was separated from the excess o-phen and a red compound of Fe chelated fromΔnifE Av1 with o-phen. An amount of Fe atom in the chelate was calculated by Lechenko and Sadkov (1984). Extraction of FeMoco was performed according to the new method of McLean et al (1989). A reconstituent solution containing Mn designated as RS-Mn was prepared as described (Huang et al, 1994). The final concentrations of KMnO4, ferric homocitrate, DT, DTT, Na2S and Hepes in RS-Mn were 0.64, 1.37, 4.96, 16.80, 1.46 and 7.52 mmol/L, respectively. Determination of protein concentration and H+-reduc-tion activity, and SDS-PAGE were carried out by Christiansen et al (1998) and Huang et al (1994; 2001; 2002). The final concentration of Na2ATP, CP (Na2.creatine phosphate) and CpK (creatine phosphokinase) in MgATP generation system are 7.34, 20.44 and 0.13 g/L, respectively. Unless described elsewhere, all of the buffer solutions were carefully degassed and filled with Ar and contained 25 mmol/L Tris (pH 7.4) and 1.7 mmol/L DT. All protein manipulations were performed under Ar. 2 RESULTS AND DISCUSSION 2.1 Purification of Δ nifE Av1 The nitrogenase in the crude extract of ΔnifE is much susceptible to heat treatment (Christiansen et al, 1998). In order to prevent the ΔnifE Av1 from a damage resulted possibly from heat treatment, the heat treatment was omitted. ΔnifE Av1 and ΔnifE Av2 could be basically separated from each other on DE52 column by developing with a NaCl gradient from 0.1 to 0.5 mol/L (Fig.1). After the chromatography on the second DE52 column, the partially purified ΔnifE Av2 (544 mg) was obtained from the crude extract (22 782 mg), and a basically pure ΔnifE Av1 (473 mg) was obtained after further purification on DE52 column, Sephacryl S-300 and Q-Sepharose columns. The analysis by SDS-PAGE shows that ΔnifE Av1 and ΔnifE Av2 were almost the same as Av1 from OP strain and Av2 from NifBmutant (UW45), respectively (Fig. 2). This indicates that ΔnifE Av1 and ΔnifE Av2 from the mutant DJ35 are similar to Av1 and Av2 from OP strain in both of the kind and molecular weight of subunits. 2.2 Activation in vitro of ΔnifE Av1 2.2.1 Activation by FeMoco FeMoco was extracted from OP Av1 adsorbed to DE52 with NMF/DNF (1:9 (V/V)) / Bu4NBr (tetrabutyl-ammonium bromide) (Mclean et al, 1989). A range of activity yield for FeMoco extracted by the method was 55%-78% (McLean et al, 1989). When NifB- Av1 (0.34 mg) and ΔnifE Av1 (0.69 mg) were incubated with 10 µL of the FeMoco, they were significantly activated; and NifB- Av1 and ΔnifE Av1 did not exhibit activity after incubation with NMF/DNF (Table 1). From the activity of the activated NifB- Av1, an activity of FeMoco extracted from 1 mg OP Av1 was calculated to be 1 822 nmol H2-formed/min-1. The activity was in the range of OP Av1(Huang et al, 2001; 2002). It indicates that the FeMoco extract is suitable for the activation of FeMocodeficient Av1. The high specific activity (2000 nmol C2H2 min-1·reduction·mg -1 protein) of NifB- Av1 activated by FeMoco was obtained only after the protein incubated with an excess amount of FeMoco was separated from NMF by chromatagraphy on DE52 column (McLean et al, 1989). The activity of NifB- Av1 incubated with FeMoco increased with an increase of FeMoco added till the added FeMoco was excess over NifB- Av1 (Shah and Brill, 1977). When incubated with the same amount of FeMoco, the specific activity of ΔnifE Av1 was about 50% of that for NifB Av1; but the amount of NifB Av1 was about 50% of that forΔnifE Av1 (Table 2). The difference in the activity between the two Apo-Av1 could be resulted from the ratio of FeMoco to Apo-Av1. It shows that the ΔnifE Av1 could be one of FeMoco-deficient Av1, which hardly exhibits any activity, and could be activated by FeMoco extracted from OP Av1 (Tal et al, 1991). 2.2.1 Activation by reconstituent solution containing Mn After ΔnifE Av1 (134 mg) treated with o-phen was chromatographied on Sephadex G-25 column, ΔnifE© Av1 (78 mg) was separated basically from a red Fe chelate (1 516 ng Fe) and an excess o-phen. Fe atoms chelated from ΔnifE Av1 was estimated to be 3-5 atoms per molecule of protein. LikeΔnifE Av1, ΔnifE© Av1 had hardly any activity when complemented with Av2. After an incubation with RS-Mn in the presence of both MgATP regeneration system and Av2,ΔnifE© Av1 was significantly activated in vitro (Table 2). But the activation did hardly happen not only in the absence ofΔnifE© Av1, but also in the absence of MgATP or Av2 or RS-Mn. This indicates that: (1) the protein activated by RS-Mn could be ΔnifE© Av1, rather than a residual Apo-Av1 in Av2; and (2) Like the full activation of ΔnifH Av1 by FeMoco (Tal et al, 1991), the activation of ΔnifE© Av1 by RS-Mn requires the simultaneous presence of both MgATP and Fe protein. Unlike ΔnifE© Av1, ΔnifE Av1 was not activated in vitro by RS-Mn in the presence of both MgATP regeneration system and Av2. As an anaerobic treatment with o-phen made OP Av1 loosen (Lechenko and Sadkov, 1984), it is reasonable to postulate that the treatment could also make ΔnifE Av1 loosen, leading to a substantial conformational rearrangements in ΔnifE Av1. Perhaps, this rearrangement is also able to be required for an assembly of metalloclusters in the clusters-deficient Av1 when the protein is incubated with this kind of reconstituent solutions in the presence of MgATP and Av2. 3 CONCLUSION ΔnifE Av1 purified from DJ35 is shown to be basically purified by SDS-PAGE analysis and one of FeMoco-defi-cient Av1. The comparison between the crude extracts from three strains showed that ΔnifE Av1 could be different from some of FeMoco-deficient Av1 in the properties (Tal et al, 1991). Therefore, its biophysical and biochemical studies including analysis of its crystalline structure are urgent, which are on the way in our laboratory. The treatment with o-phen could lead to a substantial conformational rearrangements of ΔnifE Av1, resulting in an assembly of FeMnco in the protein by incubation with RS-Mn. As Fe atoms in the chelate mainly come from P-cluster in Av1 under Ar (Lechenko and Sadkov, 1984), ΔnifE Av1 treated by o-phen under the same condition is a Apo-Av1© in which FeMoco is almost deleted and P-cluster is partially deleted either. The fact that RS-Mn was able to activate this ΔnifE Av1© indicates that both FeMnco and some of P-clusters could be assembled by the incubation of ΔnifE Av1© with RS-Mn. Therefore, this study should help us fetch some of key evidence to really characterize the nitrogenase MnFe protein. The interesting research work still goes on the way in our laboratory. REFERENCES
(Managing editor: HE Ping ) Copyright 2003 - Acta Botanica Sinica. Free, full-text also available from http://www.chineseplantscience.com The following images related to this document are available:Photo images[as03010t2.jpg] [as03010f1.jpg] [as03010t1.jpg] [as03010f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}