 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
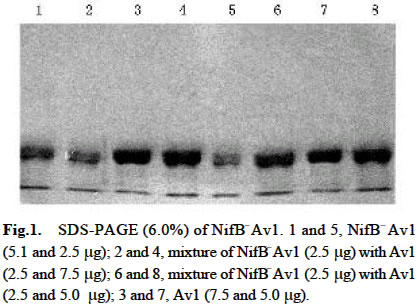
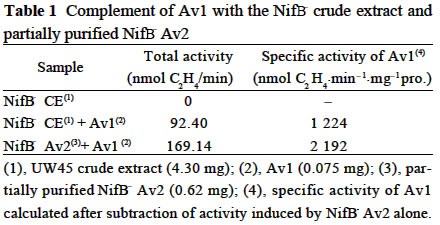
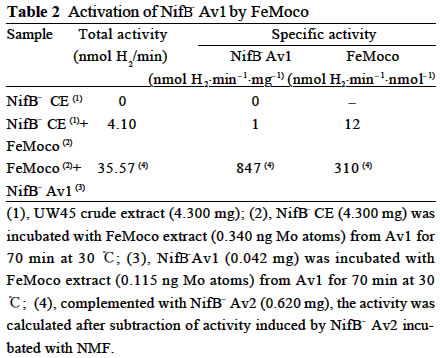
Acta Botanica Sinica, Vol. 45, No. 7, 2003, pp. 820-824 Crystallization of Nitrogenase MoFe Protein (NifB-Av1) from a nifB Mutated Strain UW45 of Azotobacter vinelandii ZHAO Ying1, L Ü Yu-Bing1, ZHAO Jian-Feng1, ZHOU Jun-Xian2, QIAN Zhong-Xing1 , WANG Yao-Ping2*, HUANG Ju-Fu1* (1. Key Laboratory of Photosynthesis and Environmental
Molecular Physiology, Institute of Botany, The
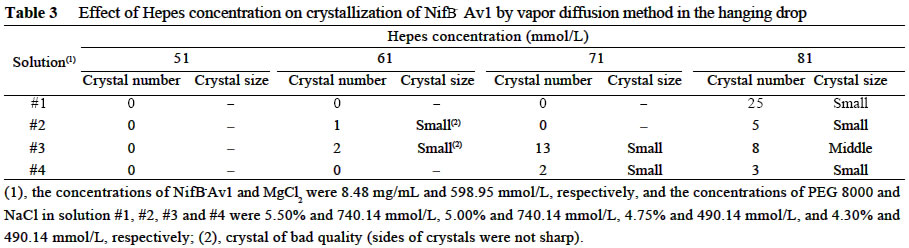
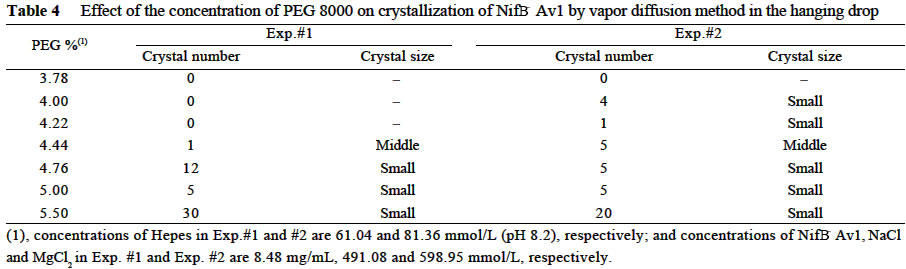
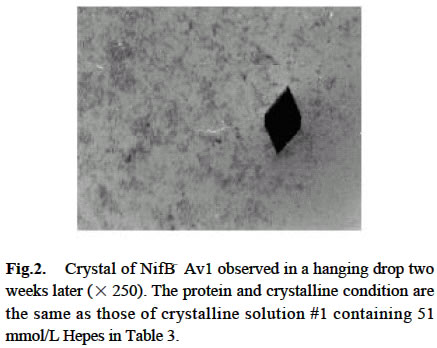
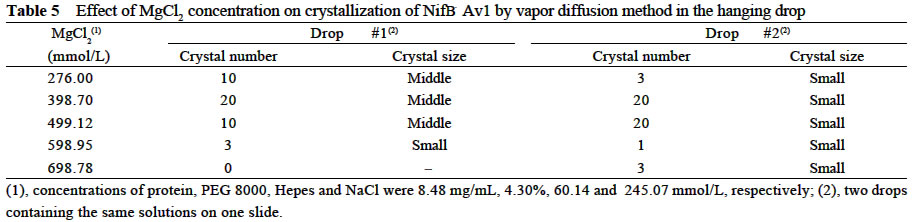
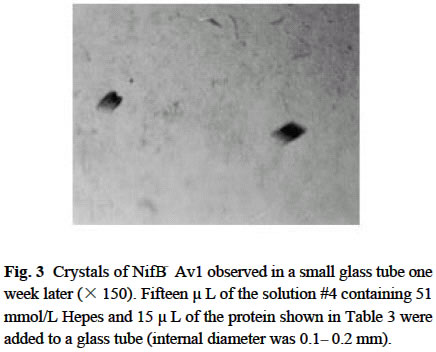
Chinese Academy of Sciences, Beijing 100093, China; Supported by the State Key Basic Research and Development Plan of China (001CB1089-06), the National Natural Science Foundation of China (30270296) and the National Manned Space Engineering Project of China. Received: 2002-05-08 Accepted: 2002-08-28 Code Number: as03011 Abstract: Six hundred and 28 mg of NifB- Av1 was obtained by a chromatography twice on DE 52 columns and Sephacryl S-300 column from the crude extract (37 677 mg) of a nifB mutated strain UW45 of Azotobacter vinelandii Lipmann. The protein was almost homogeneous as determined by Coomassie staining of SDS gels. The analysis by SDS-PAGE showed that NifB-Av1 was similar to Av1 from wild-type strain of A. vinelandii (OP) in the kinds of subunits (α and β subunit). When complemented with Av2, NifB-Av1 had hardly any H-reducing activity, but could be significantly activated by FeMoco extracted from Av1. Under a suitable condition for crystallization, short dark-brown rhombohedral crystals could be obtained from NifBAv1. Both of the longest sides of the biggest crystal were 0.1 mm. The time of the formation of crystals and number, size, quality and shape of crystals obviously depended not only on the kinds and concentrations of the components in the precipitant solution, but also on the methods for crystallization and technical bias, etc. The preliminary results showed that the crystal seemed to be formed from NifB-Av1. Key words: mutant strain UW45 of Azotobacter vinelandii ; nitrogenase NifB-Av1; crystallization Mo-containing nitrogenase is composed of two separable proteins called MoFe protein (Av1) and Fe protein (Av2). FeMoco and P-cluster in Av1 play a very important role in the structure and function of the protein (Kim and Rees, 1992). The biosynthesis and insertion of FeMoco in nitrogen fixation bacteria in vivo are controlled by nifB, nifN, nifE, nifQ, nifV, nifH and nifX (Allen et al, 1995; Rangaraj et al, 2001), and the biosynthesis of P-cluster is controlled by at least nifZ (Huang et al, 1999). Therefore, the structure and function of these nitrogenases from these genes deleted or mutated strains have been vigorously studying. Since 1977, a crude extract from UW45, which is a nifB point-mutation strain, has been used as an Apo-Av1 activated by FeMoco and its chemical model in order to study the structure and function of both nitrogenase and the model (Shah and Brill, 1977). And the protein was purified to >95% purity in 1990 (Paustian et al, 1990). Since UW45 deletes a NifB -cofactor (iron- and sulfur-containing precursor to FeMoco), it only synthesizes a FeM oco-deficient NifB-Av1 which is similar to Av1 in the structure of P-clusters and the environment of polypeptides around the clusters (Allen et al, 1995; Christiansen et al, 1998). Dislike ΔnifH Av1 from nifH deleted strain DJ54 (Tal et al, 1991), the insertion of FeMoco into NifB- Av1 in vitro is independent on an incubation with both Av2 and MgATP. A comparative study on their exact structures should help us to understand well the mechanism of reconstitution in vitro of the protein with FeMoco and its chemical model. One of the biggest and most important breakthroughs of structural studies on nitrogenase has been made after the completion of an X-ray diffraction analysis of crystals of Av1 and its complex proteins with Av2 (Kim and Rees, 1992; Schindelin et al, 1997). It is reasonable to think that the X-ray diffraction analysis of crystals of cluster-deficient Av1 from mutant strains is also of great importance for further understanding the mechanism of nitrogen fixation. However, the growth of crystal suitable for the X-ray diffraction analysis is usually a main hindrance for crystallography (McPherson ,1983). Therefore, it is necessary to screen the optimal conditions for crystallization and crystal growth of NifB -Av1. 1 MATERIALS AND METHODS Growth of mutant strain UW45 of Azotobacter vinelandii Lipmann and purification of NifB-Av1 and NifB-Av2 from the strain were carried out according to Shah and Brill (1977) and the modifie d method of Paustian et al (1990), respectively. In order to prevent the NifB -Av1 from a damage resulted possibly from heat treatment at 52 °C, the crude extract of UW was not heated. The crude extract (37 677 mg) was applied to DE52 column, followed by developing with NaCl gradient (0.1-0.5 mol/L) in 25 mmol/L Tris buffer (pH 7.4) containing 0.3 mg/mL Na2S2O4 (DT). After chromatography again on the second DE52 columns, the partially purified NifB-Av1 (1 922 mg) and NifB-Av2 (1 017 mg) were obtained. The NifB-Av1 was further purified on Sephacryl S-300 column (2.5 cm × 72.0 cm) and concentrated on a small DE52 column. Extraction of FeMoco from Av1 were performed according to the methods of Shah et al (1997) and Huang et al (1999). Determination of protein concentration and substrate-reducing activity, and SDS-PAGE were carried out according to Shah and Brill (1977) and Paustian et al (1990). Crystallization of NifB-Av1 was performed according to Drenth et al (1991) and Huang et al (1999). Unless described elsewhere, the crystals were grown by vapor diffusion using the hanging drop method for 7 d at 20 °C. All of the operations were done under Ar. All of the solutions used were rigorously degassed and filled with Ar. The concentrations of DT, Tris, and NaCl in the NifBAv1 solution were 0.3 g/L, 25 mmol/L and 250 mmol/L. The concentration of DT in precipitant solution was 12.0 mmol/L. 2 RESULTS AND DISCUSSION 2.1 Identification of NifB-Av1 The NifB-Av1 (628 mg) from the last DE52 column was basically homogeneous as determined by Coomassie staining of SDS gels, and both the positions and amount ratio of two main bands were almost the same as those of α and β subunits from Av1 (Fig. 1). And a mixture of NifB-Av1 with Av1 on the gel showed that the bands for the two proteins well overlapped each other, respectively. This indicates that kind and composition of the NifB-Av1 are almost the same as those of Av1. As reported previously in other laboratories (Shah and Brill, 1977; Christiansen et al, 1998; Huang et al, 1999), the crude extract of UW hardly exhibited any C2H2-reducing activity, but was able to be efficiently complemented with Av1 (Table 1); and either the crude extract or NifB-Av1 could be significantly reactivated by FeMoco extracted from Av1 with NMF (Table 2). The activities of the Av1 complemented with the partially purified NifB-Av2 were almost the same as that of the same batch Av1 complemented with Av2 (Huang et al, 1999). This indicates that NifB-Av2 is similar to Av2 in the activity and FeMoco can activate the inactive NifB-Av1 in vitro. 2.2 Crystallization of NifB-Av1Besides the property, purity and concentration of a protein, there are many factors affecting crystallization of the protein, such as the kind, concentration and ratios of all components in the precipitant solution, method of crystallization, pH of the solution, temperature and technical bias (Schindelin et al, 1997; McPherson, 1983; Drenth et al, 1991; Kim and Rees, 1992; Huang et al , 1999). Therefore, it is necessary to optimize the above conditions in order to obtain crystals of NifB-Av1 suitable for X-ray diffraction analysis. 2.2.1 Effect of precipitant solution on the crystallization Hepes buffer Within the Hepes concentrations shown in Tables 3 and 4 (pH 8.2), the possibility of nucleation of NifB-Av1 in the first week increased obviously with the increase of Hepes concentration. In the precipitant solution #1 containing 51 mmol/L Hepes shown in Table 3, a crystal formed in the second week could gradually grow. After incubation for four months, the longest two sizes and diagonals of the crystal were 0.10 mm × 0.10 mm and 0.17 mm×0.09 mm, respectively (Fig.2). It is indicated that the concentration of the buffer in the precipitant solution could affect the electric charge and conformation of protein, resulting in a change of the possibility and rate of nucleation. PEG 8000 Under the conditions shown in Table 4, the concentration of PEG 8000 had also a significant effect on the crystal number and size of NifB-Av1. In comparison with ΔnifZ Av1 (Huang et al, 1999), the optimal PEG concentration for crystallization of NifB-Av1 at which the crystals were bigger in size and fewer in number was slightly higher. The protein crystals could not be formed or a large amount of small crystals were formed when the PEG concentration was out of this range. PEG could absorb water from the environment around the protein molecules, resulting in decreasing protein dissolution (Drenth et al, 1991). When the PEG concentration was lower than a critical value, its crystals could not be formed. When the concentration was too high, the acceleration or disturbance of crystal formation was resulted from the acceleration of loss of water from the protein, leading to the following possibilities that the crystalline nuclei could not be formed or were too much. Only under the optimal concentration of PEG, the rate of losing water could benefit a formation of a few nuclei and their slow growth, leading to formation of the single and big crystals of good quality. MgCl2 and NaCl The concentration of MgCl2 could significantly affect the number and size of crystals from the mutant protein (Table 5). In comparison with ΔnifZ Av1 (Huang et al, 1999) , the optimal concentration (about 250 mmol/L) for its crystallization was slightly lower. Like PEG, both MgCl2 and NaCl could not only absorb water molecules from the environment around the protein, but also release cations (Mg2+ and Na+, respectively), leading to an effect of the crystalline process by a change of the electric charges on proteins. In general, the effect increases with the increase of the ion concentration and square charge number of cations (McPherson, 1983; Tal et al, 1991; Huang et al, 1999). A body of experiments in our laboratory exhibited that the effect of MgCl2 was obviously higher than that of NaCl. 2.2.2 Effect of technical bias on the crystallization Under the same conditions for crystallization by the vapor diffusion using the hanging drop method, the crystals formed in two drops sited on one slide were obviously different from each other in both the number and size, although their crystal qualities were not easy to be differentiated (Table 5). Two technical biases exhibited: (1) the error of the very small sample volume which led to a significant change in the concentration; and (2) it was possible to let the protein be differently mixed with the precipitant solution when the slide was turned over and put against the reservoir. Therefore, it could make the water molecules lose with different rates from the environment around protein molecules, leading to a different process of crystallization. 2.2.3 Effect of crystallization method on the crystallization Under the same conditions including the same protein and precipitant solutions, the results obtained by the vapor diffusion was different from those by the liquid/ liquid diffusion, and sometimes the differences were very obvious. In one week, the crystals were not formed by the vapor diffusion, but more than 10 middle crystals were observed by the liquid/liquid diffusion using the same protein and precipitant solution #4 containing 51 mmol/L Hepes shown in Table 3 (Fig.3). In comparison with the crystals shown in Fig.2, the crystals exhibited thicker. Both the rate and route for diffusion of water molecules from the environment around protein molecules could be different when the protein was crystallized by the different diffusion method, resulting in different processes for crystallization. The observation by the optical differential microscope showed that the crystals formed from the solution containing NifB-Av1 could be crystals formed from a protein, not from compounds in the precipitant solution. The dark brown color of crystals from both Av1 and ΔnifZ Av1 were related to the large amount of iron in the proteins (Huang et al,1999). Since NifB-Av1 still has P-cluster of which iron content is about a half of Av1, the color of the crystal is reasonable to be brown. Since the NifB-Av1 used for crystallization was basically homogeneous judged by SDS-PAGE, it was almost impossible that the crystals were from other contamination proteins. Finally, like ΔnifZ Av1 (Huang et al, 1999), NifB-Av1 could be crystallized only under an anaerobic condition and the formed crystals disappeared after an exposure to air. Besides the protein, only DT can be oxidized in the crystalline system while DT crystal can not be brown in color. Therefore, it seems to be reasonable to suppose that the formed crystals were those of the protein. However, the final conclusion could be made only after X-ray diffraction analysis of the crystals. The research work still goes on the way. ACKNOWLEDGEMENTS: The authors wish to thank Mr. ZHANG Hua-Feng for his help in some experiments. REFERENCES:
(Managing editor: HE Ping) Copyright 2003 - Acta Botanica Sinica. Free, full-text also available from http://www.chineseplantscience.com The following images related to this document are available:Photo images[as03011t2.jpg] [as03011f2.jpg] [as03011t4.jpg] [as03011f1.jpg] [as03011t5.jpg] [as03011f3.jpg] [as03011t3.jpg] [as03011t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}