 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
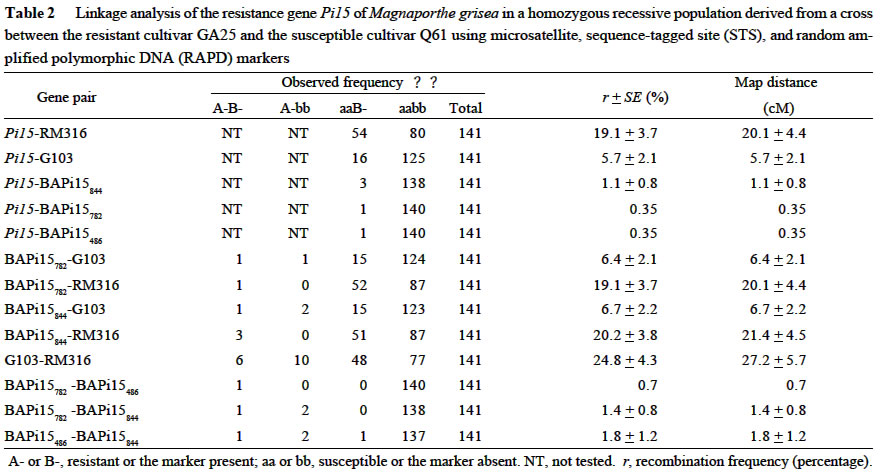
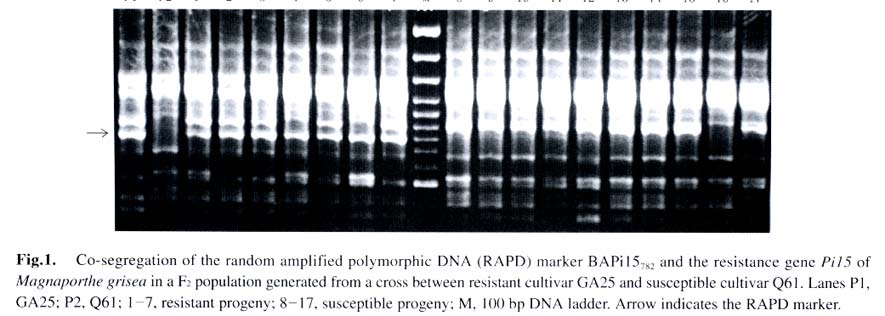
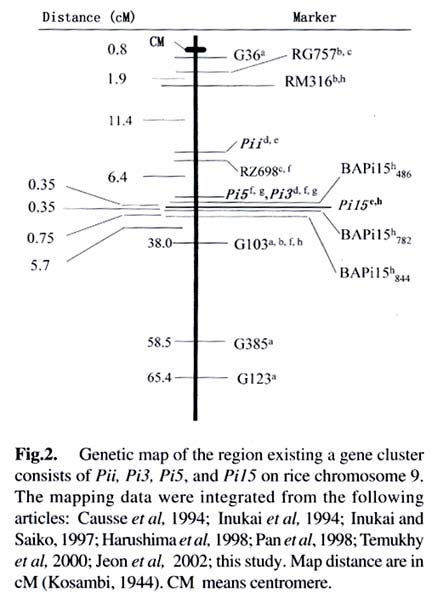
Acta Botanica Sinica, Vol. 45, No. 7, 2003, pp. 871-877 Fine Mapping of the Blast Resistance Gene Pi15, Linked to Pii, on Rice Chromosome 9 PAN Qing-Hua1*, HU Zhen-Di1, Tanisaka TAKATOSHI2, WANG Ling1 (1. Laboratory of Plant Resistance and Genetics, College of Resources and Environment, South China Agricultural University, Guangzhou 510642, China; * Author for correspondence. E-mail: Received: 2003-03-05 Accepted: 2003-04-21 Code Number: as03016 ABSTRACT The gene Pi15 for resistance of rice to Magnaporthe grisea was previously identified as being linked to the gene Pii. However, there is a debate on the chromosomal position of the Pii gene, because it was originally mapped on chromosome 6, but recent work showedit might be located on chromosome 9. To determine the chromosomal location of the Pi15 gene, a linkage analysis using molecular markers was performed in a F2 mapping population consisting of 15 resistant and 141 susceptible plants through bulked-segregant analysis (BSA) in combination with recessive-class analysis (RCA). Out of 20 microsatellite markers mapped on chromosomes 6 and 9 tested, only one marker, RM316 on chromosome 9, was found to have a linkage with the Pi15 gene with a recombination frequency of (19.1 ±3.7)%. To confirm this finding, four sequence-tagged site (STS) markers mapped on chromosome 9 were tested. The results suggested that marker G103 was linked to the Pi15 gene with a recombination frequency of (5.7 ±2.1)%. To find marker(s) more closely linked to the Pi15 gene, random amplified polymorphic DNA (RAPD) analysis was performed. Out of 1 000 primers tested, three RAPDmarkers, BAPi15486,BAPi15782 and BAPi15844 were found to tightly flank the Pi15 gene with recombination frequencies of 0.35%, 0.35% and 1.1%, respectively. These three RAPD markers should be viewed as the starting points for marker-aided gene pyramiding and cloning. A new gene cluster of rice blast resistance on chromosome 9 was also discussed. Key words: bulked-segregant analysis (BSA) ; microsatellite; rice (Oryza sativa); Magnaporthe grisea ; RAPD; recessive-class analysis (RCA); sequencetagged site (STS) Resistance to the rice blast fungal pathogen (Magnaporthe grisea) is of major importance to rice breeding programs in many countries (Ou, 1985). More than 40 major resistance genes have been reported (Imbe et al, 1997; Hayashi et al, 1998; Pan et al, 1999; Zhuang et al, 1999; Tabien et al, 2000; Fujii et al, 2000; Fukuoka and Okuno, 2001). Because the spectrum of virulence is dynamic in field populations, genes for resistance, especially single major genes, quickly succumb to new virulent races of the pathogen (Yamasaki and Niizeki, 1963; Ou, 1979; Kiyosawa, 1989; Ling et al, 1990; Chen et al, 2001). Addressing possible solutions for the breakdown of resistance, Kiyosawa (1982) critically reviewed the utilization of resistance genes in breeding programs. They are, multilines, deployment of resistant cultivars, rotation of resistant cultivars; mixed cultivation of resistant cultivars, and pyramiding several major and/or minor resistant genes into single elite cultivars. As for the last strategy, the field experience and computer simulations indicated that the resistance of cultivars which carry several resistance genes should be longer-lasting than those of cultivars carrying single resistance genes (Kiyosawa, 1982). However, gene pyramiding has been impaired by the conventional inoculation method, because no universal set of isolates that can completely distinguish reactions of the individual resistance genes exists (McCouch et al, 1994; Miyamoto et al, 1996; Naqvi and Chattoo 1996; Huang et al, 1997). Molecular marker technology is a certain approach to clear this difficulty, because the presence of flanking markers tightly linked to resistance genes will allow selection and maintenance of the desirable resistant genotypes in breeding process (Hittalmani et al, 1995; Naqvi and Chattoo, 1996; Huang et al, 1997). Further markers tightly linked to the target resistance gene should be as an essential starting point to clone the gene through map-based approach. In the previous study, the authors identified the Pi15 gene through allelism tests with Japanese differential cultivars, and further predicted that this gene linked to the known resistance gene Pii (Pan et al, 1998). This locus was originally mapped on chromosome 6 (Kiyosawa, 1967). But recent evidence suggests that Pii might locate on chromosome 9 (Inukai et al, 1994; Inukai and Saiko, 1997; Pan et al, 1998). The objective of this research was to determine the chromosomal position of the Pi15 gene, and to identify molecular markers tightly linked to the resistance gene for subsequent marker-aided gene pyramiding and cloning. 1 MATERIALS AND METHODS 1.1 Plant materials As for gene mapping, the Pi15 donor cultivar GA25 was crossed to an indica susceptible cultivar Q61, and their F2 population was used as a mapping population. For the resistance spectrum tests, two cultivars Fujisaka 5 and C104PKT, both carry resistance genes at the loci in the vicinity of the Pi15 locus, as well as five cultivars, Fukunishiki, Toride 1, C101A51, PiNo. 4 and Tsuyuake, which possess broad-spectrum resistance to isolates originated from Japan, Thailand and China, were used (Table 1). All the rice seeds were pregerminated by soaking in water at 25 °C for 48 h, and sown in a plastic pot (34 files × 15 rows) filled with black organic soil treated with fungicide solution. Each pot was placed in a plastic tray (58 cm×38 cm×8 cm). To construct the mapping population, the parental cultivars and a susceptible check cultivar Sariceltik were sown at both ends of the F2 population in a tray. To test the resistance spectrums, the eight cultivars selected were randomly sown with other 60 cultivars including Sariceltik in a tray, and each cultivar was sown five seeds in a row. Seedlings were grown in a greenhouse at 20-35 °C for about three weeks before inoculation. 1.2 Conidial inoculation and disease scoring M. grisea isolate CHL0416 that is avirulent on GA25 and virulent on Q61 was selected for constructing the mapping population. Nine additional isolates those possess clear infection types on the cultivars selected were used for the resistance spectrum tests (Table 1). Inoculum was prepared from cultures grown on rice polish agar medium (20 g of rice polish powder, 2 g of yeast extract, and 11 g of agar per liter of distilled water). Two pieces of sterile filter paper discs were placed onto the medium in a 10 cm glass petri plate, and incubated at 25 °C for about 10 d until the surface of the plate was almost covered with mycelial growth. The growth was gently scraped in sterilized distilled water with a metal spatula on a clean bench, and then the plate was placed on the sporulation incubator under dark blue light for 2-3 d to facilitate condition of the culture. The culture was flooded with distilled water, and conidia were released by scraping. The suspension was filtered through gauze mesh, and 0.5% solution of gelatin (DGF STOESS, Germany) was added as a surfactant. The concentrations of conidia ranged from 1 × 105 to 5 × 105 conidia/mL. At the four- to six-leaf stage, seedlings were transferred into an inoculation incubator, and inoculated by spraying 70-80 mL of spore suspension per tray. After inoculation, the seedlings were kept in the inoculation incubator at 25 °C with saturated humidity for 20 h, and then transferred to a moist vinyl-tunnel at 25-30 °C. Disease reactions were scored for about 7 d after inoculation by use of a 6-class scale described by Pan et al (1996). For data analysis, the seedlings of classes 0-2 were referred to as resistant, and those of classes 3-5 were referred to as susceptible. 1.3 DNA extraction and amplification Total DNA was extracted from frozen leaves of rice plant according to Doyle and Doyle (1990). Amplification reactions were performed with the Programmable Thermal Controller PTC 100TM (MJ Research Inc. Watertown, Massachusetts) or the GeneAmp ® PCR System 9700 (PE Applied Biosystems, Foster City, California). For microsatellite and sequence-tagged site (STS) analyses, primer sets of microsatellite (Temnykh et al, 2000) and STS (Inoue et al, 1994) were synthesized by Shanghai Sangon Biotechnology Co., Ltd. (Shanghai, China) and Shanghai BioAsia Biotechnology Co., Ltd. (Shanghai, China), respectively. The PCR procedures described by Temnykh et al (2000) and Inoue et al (1994), respectively, were followed with the exception that both PCR products were separated on 3.3% polyacrylamide denaturing gels. For RAPD analysis, random 10-mer primers were purchased from Shanhai BioAsia Biotechnology Co., Ltd. Each reaction (20 µL) contained 0.16 µL of Taq DNA polymerase (5 unit/µL, GeneTek), 2 µL of 10 × reaction buffer mix (GeneTek), 0.4 µL of dNTPs (10 mmol/L), 1.6 µL of primer (2.5 µmol/L), 0.8 µL of template DNA (10-20 ng/µL) and 15.04 µL of ddH2O. The amplification was programmed for 45 cycles of 1 min at 94 °C, 1 min at 35 °C and 2 min at 72 °C. The reaction products were then visualized by electrophoresis on 1.2% agarose gels containing ethidium bromide in 1 × Tris-bo-rate-EDTA (pH 8.0). 1.4 Identification of linkage marker The bulked segregant analysis (BSA; Michelmore et al, 1991) was used to identify candidate markers linked to the resistance locus. Two contrasting bulks were prepared, each containing DNA from 15 resistant or susceptible F2 individuals. Candidate markers were confirmed by the recessive-class analysis (RCA; Zhang et al, 1994) with each individual of the bulks and 126 additional susceptible individuals selected from the F2 population. RCA was chosen for verification because of the reliable reactions (diagnostic symptoms) and high efficiency to locate the gene on its chromosome (Zhang et al, 1994; Lin et al, 1996; Miyamoto et al, 1996). By assuming that all 141 susceptible plants were homozygous for the recessive allele at the target Pi15 locus, the recombination frequency (rRM) between a marker and the resistance gene was calculated by the maximum likelihood method (Allard, 1956), based on the molecular data from the 141 susceptible plants: rRM = NaB / 2NT, in which NT is the total number of the susceptible individuals tested, NaB is the number of the susceptible individuals presenting a marker (Table 2). Likewise, the recombination frequencies (rMM) among the marker linked to the resistance gene were estimated by the balance: rMM = 2NAB + NAb + NaB / 2NT, in which NAB, NAb , and NaB are the numbers of the susceptible individuals presenting markers A and B, A only, and B only, respectively (Table 2). The variance is given by Vr = 1 / NTIr, in which Ir is supplied according to Allard's Table 8 (Allard, 1956). The recombination frequency was transformed into centimorgans (cM) according to the Kosambi function (Kosambi, 1944). 2 RESULTS 2.1 Developing the mapping population A total of 1 314 F2 progenies derived from the cross of GA25 with Q61 were inoculated with isolate CHL0416, which is avirulent to GA25 and virulent to Q61. Segregation of resistant and susceptible progenies fitted a 3 : 1 ratio in the F2 population (979R : 335S, Χ2 = 0.15, P > 0.70), consistent with the hypothesis that GA25 carries a single dominant locus conferring resistance. Since many susceptible individuals did not survived when they were transplanted in pots, 15 resistant and susceptible plants were selected for BSA, and 141 susceptible individuals including 15 susceptible plants used in BSA were subjected to RCA. 2.2 Confirming the map location of Pi15 locus with microsatellite and STS markers To determine whether the Pi15 gene is located on chromosome 6, the initial analysis was focused on the microsatelite markers around the former position of the Pii gene (Kiyosawa, 1967; Temnykh et al , 2000). Ten microsatelite markers, RM121, RM136, RM3, RM162, RM275, RM30, RM340, RM103, and RM141 were tested, and no markers were found to have a linkage with the Pi15 gene (data not shown). The second analysis was focused on 10 microsatelite markers surrounding the position of Pii on chromosome 9 predicted in the previous study (Pan et al, 1998). They are RM285, RM316, RM219, RM105, RM321, RM257, RM242, RM108, RM288 (Temnykh et al, 2000), and OSR28 (Akagi et al, 1996). Two markers, RM316 and RM257, were considered as candidate markers through BSA, and only RM316 was verified to have a linkage with the Pi15 gene with a recombination frequency of (19.1 + 3.7) % through RCA (Table 2). To find additional markers flanking the Pi15 gene, four STS markers, G36, G103, G385 and G123 mapped on chromosome 9 (Inoue et al, 1994; Harushima et al, 1998) were tested. The results indicated that only G103 was linked to the Pi15 gene with a recombination frequency of (5.7 + 2.1)% (Table 2). 2.3 Fine mapping the Pi15 gene with RAPD markers To find markers tightly linked with Pi15 gene, RAPD analyses were performed with the same process of the microsatellite and STS marker assays. A total of 1 000 random primers were tested, and three markers, designed as BAPi15486, BAPi15782 and BAPi15844 were confirmed to display tightly linkage to the Pi15 gene with recombination frequencies of 0.35%, 0.35% and 1.1%, respectively (Table 2). Cosegregation of the RAPD marker BAPi15782 and the resistance gene Pi15 are shown in Fig. 1. The genetic map of the Pi15 gene and other resistance genes, Pii, Pi3 and Pi5 mapped on chromosome 9 are shown in Fig. 2. In this comprehensive map, the positions of anchor markers, G36, G103, G385 and G123, RG757 and RM316, and RZ698 are integrated from the maps constructed by Harushima et al (1998), Temnykh et al (2000), and Causse et al (1994), respectively. The position of the resistance gene Pii is estimated by the recombination frequencies of Pii with Pi3 (Inukai et al, 1994) and Pi15 (Pan et al, 1998). Furthermore, the map location of Pi3 is determined on the recombination frequencies of the resistance gene with the anchor markers RZ689 and G103 (Inukai et al, 1997). As to the position of the resistance gene Pi5, it is estimated at the same position with Pi3, because recent work suggested that both resistance genes are in fact one gene (Jeon et al, 2002). 2.4 Further confirming the Pi15 gene identity through the resistance spectrum tests To find further evidence for confirming whether the Pi15 is identical to other resistance genes mapped on the loci in the vicinity of the Pi15, the resistance spectrum comparisons were performed among the donor cultivars, GA25, Fujisaka 5 and C104PKT, those carry resistance genes,Pi15, Pii, and Pi3, respectively. The different resistance spectrums mediated by 10 isolates selected from Japan, Thailand and China were observed among the three cultivars, demonstrating that the Pi15 gene is clearly distinct from the resistance genes Pii and Pi3 (Table 1). To gain further information about the potential usefulness of the Pi15 gene in rice improvement, the resistance spectrum comparisons were carried out among the Pi15 donor cultivar, GA25, and five cultivars, Fukunishiki, Toride 1, C101A51, PiNo. 4, and Tsuyuake those express resistance to many isolates existing in Japan, Thailand and China (data not shown). Complementary reactions to the 10 isolates selected were observed between the Pi15 donor cultivar GA25 and the other five cultivars, indicating that the Pi15 gene should be an excellent resistance source for improving rice cultivars with stable resistance to blast fungus pathogen (Table 1). 3 DISCUSSION The Pi15 gene was identified to display linkage with the Pii gene in previous study (Pan et al, 1998). However, there was debate on the chromosomal location of the Pii gene. Although Kiyosawa (1967) identified a linkage relationship between the Pii gene and the Piz gene on chromosome 6 with a recombination frequency of 30.9%, later works have shown that Pii is not on chromosome 6 (Goto et al, 1981; Iseu, 1992; 1993; Pan et al, 1996). On the other hand, Inukai et al (1994) first reported that a new resistance gene Pi3 is closely linked to Pii with a recombination frequency of (7.0 + 2.6)%. Furthermore, Inukai and Saiko (1997) first obtained the molecular evidence that the Pi5 gene, which is allelic or closely linked to the Pi3, was mapped at a locus flanking molecular markers RZ689 and G103 on chromosome 9. Later, the authors also identified a new resistance gene Pi15 that is linked to Pii with a recombination frequency of (15.1 + 2.8)%, and further suggested that a new gene cluster that consists of Pii, Pi3, Pi5 and Pi15 may located on rice chromosome 9 (Pan et al, 1998). Moreover, Jeon et al (2002) compared the genomic regions for lines containing Pi3 and Pi5, and concluded that both resistance genes are likely the same, because the Pi3 physical region is exactly identical to that of Pi5. Compared with the reaction patterns between the resistance donors GA25 and C104PKT (Table 1), as well as the genetic distances between the anchor marker G103 to Pi5 (3.5 cM, Jeon J S, personal communication) and Pi15 (Table 2), it clearly suggested that Pi15 is not identical, but tightly linked to Pi5 (Pi3). In order to identify markers linked to resistance genes, Michelmore et al (1991) proposed a rapid method to detect markers in the target genomic regions through BSA approach. Following this, Zhang et al (1994) proposed the more cost-effective approach, RCA, which is particularly useful for mapping genes controlling expression of a complex trait. Lin et al (1996) subsequently applied this approach in mapping the chromosomal location of the gene Xa22 conditioning resistance for rice bacterial blight disease. The results of this study provided further suggestions that combination of BSA with RCA approaches may be especially useful for mapping resistance genes, because plants with the susceptible phenotype may be clearly identified whereas plants, which escape infection may be mis-classi-fied as resistant even though they are in fact homozygous for the susceptibility allele. Moreover, some susceptible plants, which were heavily infected might not be survived for subsequent gene mapping. The resistance gene Pi15 conditions reactions complementary to those mediated by Piz, Pizt, Piz5, Pita2 and Pik m to isolates selected from Japan, Thailand and China (Table 1). As an application of it to a breeding program in Guangdong Province, China, we are undertaking to accumulate the Pi15 gene with other resistance genes into elite cultivars through marker-aided selection (Huanget al, 1997). For the longer term of cloning the Pi15 gene, we are constructing a high-resolution map of this gene in a large mapping population through the approaches mentioned above, and/or yeast artificial chromosome (YAC) clone-based subcloning approach (Monna et al, 1997; Wang et al, 1999). REFERENCES
(Managing editor: ZHAO Li-Hui) Copyright 2003 - Acta Botanica Sinica. Free, full-text also available from http://www.chineseplantscience.com The following images related to this document are available:Photo images[as03016t1.jpg] [as03016f2.jpg] [as03016f1.jpg] [as03016t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}