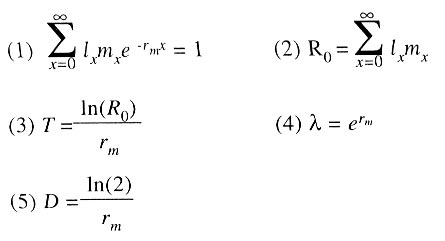|
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Agricultura Técnica, Vol. 63, No. 1, Jan - March, 2003, pp. 10-14 INCREMENTO POBLACIONAL DE Myzus persicae (Sulzer) SOBRE TRES CRUCÍFERAS HORTÍCOLAS EN LABORATORIO1 Population increase of Myzus persicae (Sulzer) on three cruciferous plants in laboratory1 Araceli Vasicek2, Francisco LaRossa3, Andrea Paglioni2 y Luis Fostel2 1 Recepción de originales:
15 de noviembre de 2001. Code Number: at03002 ABSTRACT Cohorts of Myzus persicae (Sulzer) were reared on Raphanus sativus, Brassica oleracea var. capitata and B. oleracea var. italica at 10 ± 1ºC, 90% relative humidity and a 12:12 h photoperiod. Under these conditions significant statistical differences were found between intrinsic growth rates (rm), which reached a maximum of 0.082, 0.045 and 0.028 females female-1 d-1 on R. sativus, B. oleraceea var. italica and B. oleracea var. capitata, respectively, and net reproductive rates (Ro) were 26, 6 and 3.4 females female-1 generation-1, respectively. M. persicae needs ca. around 8 days to duplicate numbers (D) on R. sativus and 16 and 30 days on B. oleracea var. italica and B. oleracea var. capitata, respectively. Results indicated this the aphid could reach greater population levels on R. sativus than on the other cruciferous plants during the fall and winter seasons, and an intercrop cultivation with other cruciferous plants could be recommended if it is the main crop. On the other hand, R. sativus is a winter weed for other vegetable crops, and thus early control could reduce posterior dispersion of the aphid. Key words: aphididae, life tables, intrinsic rate of growth, net reproductive rate, Raphanus sp., Brassica oleracea. RESUMEN Se criaron cohortes de Myzus persicae (Sulzer) sobre Raphanus sativus, Brassica oleracea var. capitata y B. oleracea var. italica a 10 ± 1ºC, 90% de humedad relativa y fotoperíodo 12:12 h. Bajo esas condiciones se produjeron diferencias significativas entre las tasas intrínsecas de crecimiento (rm), que alcanzaron un máximo de 0,082; 0,045 y 0,028 hembras hembra-1 d-1 sobre R. sativus, B. oleracea var. italica y B. oleracea var. capitata, respectivamente; las tasas netas de reproducción (Ro) fueron 26; 6; y 3,4 hembras hembra-1 generación-1, respectivamente. Este áfido necesita alrededor de 8 días para duplicarse en número (D) sobre R. sativus, y 16 y 30 días sobre B. oleracea var. italica y B. oleracea var. capitata, respectivamente. Los resultados indicaron que el áfido podría alcanzar niveles poblacionales mayores sobre R. sativus que en las otras crucíferas durante la temporada otoño-invernal, y podrían recomendarse intersiembras con otras crucíferas si el primero es el cultivo principal. Por otra parte, R. sativus es una maleza invernal en otros cultivos hortícolas, por lo cual un control temprano de la misma podría reducir la posterior dispersión del áfido. Palabras clave: aphididae, tablas de vida, tasa de crecimiento intrínseco, tasa neta de reproducción, Raphanus sp., Brassica oleracea. INTRODUCCIÓN El pulgón verde del duraznero (Myzus persicae Sulzer) es sin dudas, el áfido de mayor predominio y peligrosidad en el mundo, debido a su frecuente aparición y a la magnitud de los daños que ocasiona, tanto directos como indirectos, por la transmisión de enfermedades virales (Mier Durante, 1978). Además de producir enrulamiento y deformación de distintos órganos, esta especie es considerada por diversos autores (Van Harten, 1983; Sigvald, 1984; Spak, 1992; Castle y Berger, 1993; Syller, 1994) como el más eficiente vector de virus. Actualmente está distribuida en todo el mundo (Blackman e Eastop, 1985), y su polifagia le permite producir grandes poblaciones de individuos alados que visitan y colonizan distintos cultivos. En Argentina está muy difundida principalmente sobre hortalizas y frutales del género Prunus, sobre los cuales se comprobó el ciclo sexuado en la Provincia de Mendoza, pero no se ha verificado el holociclo en otras zonas del país (Delfino, 1983). M. persicae puede invernar en forma partenogenética sobre hospederos secundarios tanto silvestres como cultivados en lugares donde las temperaturas invernales lo permiten (Ortego y Carrillo, 1995). Esta forma de reproducción (anholociclo) se ve favorecida en gran medida por el cultivo en invernadero en el cinturón hortícola de La Plata y otras ciudades, dando lugar a sucesivas generaciones durante todo el año, casi sin interrupciones. Dada su polifagia, una gran cantidad de plantas, entre ellas crucíferas, albergan poblaciones poco numerosas de áfidos que se reproducen durante la temporada otoño-invierno y desde las cuales pueden expandirse y colonizar otros vegetales en la siguiente estación. Los parámetros biológicos y los principales estadísticos vitales de una población de insectos plaga, estimados a partir de tablas de vida desarrolladas en laboratorio, son una herramienta básica para elaborar estrategias de control (Southwood, 1994). Estas estimaciones han sido utilizadas para evaluar resistencia en plantas (Trichilo y Leigh, 1985) y como patrón para seleccionar enemigos naturales (Jansen y Sabelis, 1992). Actualmente, el control de estos insectos plaga se lleva a cabo mediante la aplicación de insecticidas químicos, lo cual eleva el costo de producción, contamina el ambiente y puede generar incompatibilidad entre el intervalo de cosecha y el período de carencia del insecticida. Los conocimientos sobre la demografía de estos insectos ayudan en la formulación de alternativas de manejo. En consecuencia, el objetivo del presente trabajo fue obtener los principales parámetros, tales como tasa neta de reproducción , tiempo generacional medio, y tasa intrínseca de crecimiento natural, para caracterizar las poblaciones otoño-invernales de M. persicae sobre tres especies de crucíferas cultivadas. MATERIALES Y MÉTODOS El presente trabajo se efectuó en el Insectario de la Cátedra de Zoología Agrícola, Facultad de Ciencias Agrarias y Forestales, Universidad Nacional de La Plata, Buenos Aires, Argentina. Las colonias madres de M. persicae provinieron de huertas comerciales de la zona (34º58' lat. Sur; 57º54' long. Oeste). Las plántulas se obtuvieron a partir de semillas puestas a germinar en un sustrato conteniendo suelo mezclado con 1/3 de compuesto orgánico esterilizado en autoclave a 120°C, 1 atm, durante 1 h, repitiendo este procedimiento a las 48 h (Dhingra y Sinclair, 1985). Sobre las mismas se transfirió una hembra adulta, la que se dejó producir ninfas durante 24 h. Luego de este período se retiraron todos los individuos recién nacidos menos uno, obteniéndose cohortes de aproximadamente la misma edad. Se aislaron ninfas neonatas, colocándolas individualmente sobre plantines de 2-4 hojas verdaderas, de rabanito (Raphanus sativus L.) cv. Gigante Mantecoso, repollo (Brassica oleracea var. capitata L.) cv. Ruby Perfection y bróccoli (Brassica oleracea var. italica L.) cv. Pinnacle. Los plantines se renovaban cada 3-4 días. La crianza individual se llevó a cabo en recipientes de plástico de 60 cm3 con agua, cerrados con una tapa perforada en el centro para la introducción del tallo. El material se protegió con envases de características idénticas con malla de tul como en la tapa para permitir la aireación. Se colocaron en una cámara refrigerada a 10 ± 1º C y humedad relativa de 60-75%. En cada crucífera se criaron simultáneamente seis cohortes de 35 individuos iniciales, totalizando 210 áfidos. Diariamente se registraron los cambios de estadio, el número de individuos muertos, y los nacimientos una vez alcanzado el estado adulto. A partir de la confección de tablas de vida se estimaron los estadísticos vitales, supervivencia por edades (lx), fecundidad por edades (mx), y los siguientes parámetros poblacionales: tasa neta de reproducción (Ro, número de hembras recién nacidas por hembra), tasa intrínseca de crecimiento natural (rm, número de hembras por hembra por unidad de tiempo), tiempo generacional medio (T); tasa finita de incremento (λ , número de veces que la población se multiplica sobre sí misma por unidad de tiempo); y tiempo de duplicación (D, número de unidades de tiempo requerido por la población para duplicarse en número) (Southwood, 1994), cuyas fórmulas son las siguientes:
donde: lx = proporción de hembras sobrevivientes a la edad x; mx = número medio de progenie hembra por hembra aún viva a la edad x; e = base de los logaritmos naturales = 2,718281.....; ln = logaritmo natural. El parámetro rm se calculó mediante sucesivas iteraciones de la ecuación 1. Mediante la aplicación del método "Jackknife" se calcularon estimadores de la rm, intervalos de confianza al 95% y los correspondientes errores estándar, con los cuales es posible efectuar comparaciones entre las cohortes (Hulting et al., 1990). Los estadísticos fueron comparados mediante la ecuación 6:
Con los parámetros obtenidos es posible modelar curvas teóricas de incremento poblacional en función del tiempo a partir de un muestreo inicial sobre el cultivo mediante la ecuación 7. (7) Nt = N0ermt donde: Nt = número total de áfidos en el tiempo t, N0 = número inicial de áfidos, e = base de los logaritmos naturales = 2,718281......; t = tiempo en días. RESULTADOS Y DISCUSIÓN En este trabajo se evaluó el comportamiento de M. persicae sobre tres crucíferas importantes cultivadas en el cinturón hortícola platense. La comparación de los parámetros demográficos (Cuadro 1) permitió caracterizar el efecto del hospedante sobre dicho comportamiento. Así, la mayor tasa intrínseca de incremento natural (rm) ocurrió sobre rabanito, mientras que en términos comparativos sobre bróccoli y repollo resultó dos y tres veces menor, respectivamente. Los restantes parámetros presentaron la misma tendencia, excepto el tiempo generacional medio (T), en el cual no se observaron diferencias significativas para las seis cohortes estudiadas. En una especie de reproducción continua, este parámetro se interpreta como la edad a la cual, si todo el esfuerzo reproductivo estuviera concentrado en ella, la tasa de reproducción neta o tasa de reemplazo (R0) sería la misma que con el esfuerzo reproductivo repartido entre varias edades (Rabinovich, 1980). Cuadro 1. Parámetros demográficos
de Myzus persicae (Sulz.) sobre rabanito (Raphanus sativus L.),
bróccoli (Brassica oleracea var. italica L.) y repollo
(Brassica oleracea var. capitata L.).
C1 y C2: cohortes 1 y 2. Se infiere que el T estaría influenciado por la temperatura, independientemente de la especie y/o variedad de hospedante perteneciente a una misma familia botánica. De Loach (1974), trabajando con repollos de la variedad Globe a 10ºC, encontró un T de 39,25 días, muy similar al obtenido aquí. En cambio, sobre tres variedades distintas de lechuga (Lactuca sativa L.), planta de otra familia, el tiempo generacional a 20ºC resultó similar, aunque rm fue sustancialmente diferente (Ricci et al., 1999). Dado que rm es un parámetro que puede resumir las características vitales de un áfido y resulta muy útil para comparar individuos (o cohortes) de una o varias especies bajo diferentes condiciones (Dixon, 1987), se puede advertir que M. persicae expresa mejor su potencial biótico sobre rabanito que en las otras crucíferas estudiadas. Por otra parte, esto también explicaría una mayor adaptabilidad sobre este hospedero, el cual crece de manera espontánea durante el invierno, le sirve de refugio, y al mismo tiempo le permite multiplicarse a una tasa aceptable. En ausencia de factores extrínsecos de mortalidad, partiendo de un número inicial de 10 hembras de cualquier edad por unidad de muestreo, empleando la ecuación 7, es posible inferir que la población se incrementaría unas 50 veces en 48-49 días sobre rabanito a 10ºC, en tanto que sobre las otras crucíferas no llegarían a 100 individuos en el mismo período. De ello se desprende que en caso de cultivarse rabanito, deben realizarse muestreos frecuentes con el fin de detectar cualquier infestación temprana y poder controlar un posible incremento poblacional. Asimismo, sería recomendable efectuar intersiembras con repollo y/o bróccoli. Si el rabanito se presenta como maleza, su eliminación temprana durante el invierno contribuiría a mantener bajas las poblaciones del áfido, beneficiando así a otros cultivos hortícolas posteriores en primavera y verano. CONCLUSIONES
LITERATURA CITADA
Full-text article also available online at http://www.inia.cl/at/agritec.htm Copyright 2003 - Instituto de Investigaciones Agropecuarias, INIA (Chile). | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||