 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
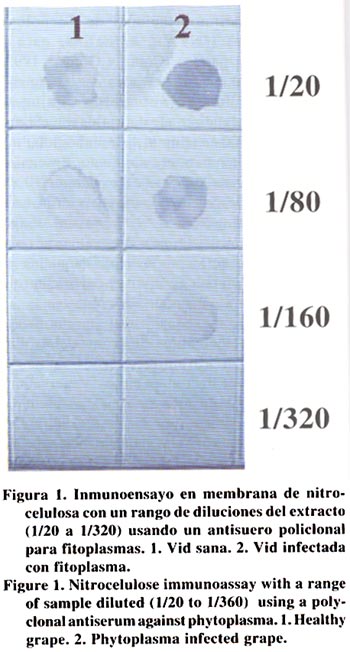
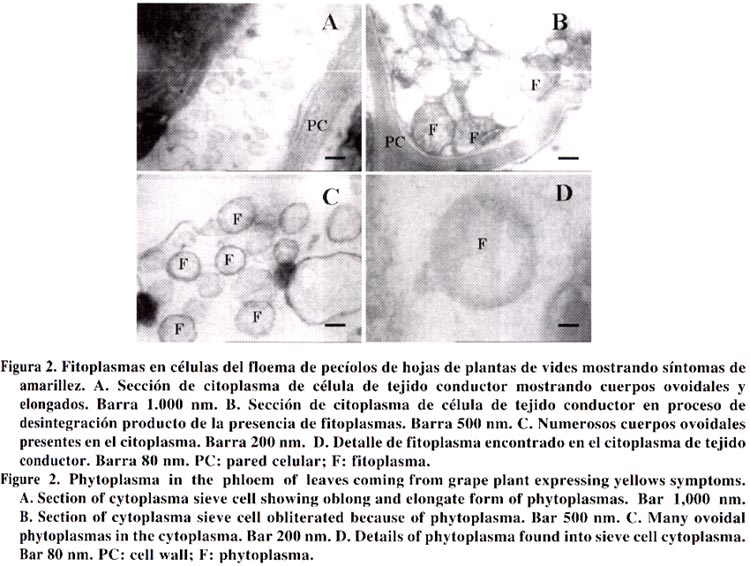
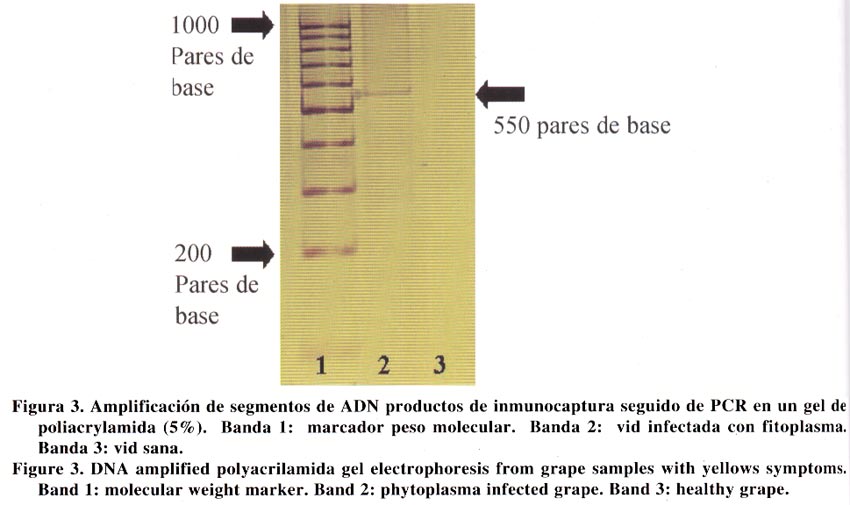
Agricultura Técnica, Vol. 63, No. 1, Jan - March, 2003, pp. 15-22 EVIDENCIAS INMUNOLÓGICAS, MICROSCÓPICAS Y MOLECULARES DE LA PRESENCIA DE FITOPLASMAS EN VIDES1 Immunological, microscopical and molecular evidence of phytoplasma in grapes1 Guido Herrera M.2* y Mónica Madariaga V.2 1 Recepción de originales:
14 de noviembre de 2001. Code Number: at03003 ABSTRACT The causal agent of grapevine yellows is associated to phytoplasm. The symptoms include leaf yellowing, rolling and abscission together to progressive vegetative decline. This study was conducted to determine whether grape (Vitis vinifera L.) plants showing yellowing symptoms were infected by phytoplasma. Grape plants from the Central zone of Chile were received in the Plant Virus Laboratory of the Centro Regional de Investigación La Platina, Instituto de Investigaciones Agropecuarias (INIA), were tested by nitrocelulose-ELISA (nc-ELISA), transmission electron microscopy (TEM) and polimerase chain reaction (PCR). Petiole and leaf vein samples coming from affected plants showed phytoplasma under transmission electron microscopy and strongly reacted with a policlonal antiserum raised against phytoplasma. Using universal primers to phytoplasma a 550 base pair DNA fragment was amplified by PCR. The evidences confirm the presence of phytoplasm in the affected grape plants. Key words: Vitis vinifera L., polimerase chain reaction (PCR) RESUMEN El agente causal de la "amarillez de la vid" es asociado a fitoplasma. Los síntomas incluyen amarillez, enrollamiento y abscisión de hojas junto a un progresivo declinamiento. Este estudio se realizó para determinar la presencia de fitoplasmas en plantas de vides (Vitis vinifera L.) afectadas con síntomas de amarillez. En el laboratorio de virología del Centro Regional de Investigación La Platina, de Instituto de Investigaciones Agropecuarias (INIA), se recibieron plantas de vides provenientes de la zona central de Chile mostrando síntomas de amarillez. Pecíolos y nervadura central de hojas se sometieron a las pruebas de nitrocelulosa-ELISA (nc-ELISA), observación de cortes finos bajo el microscopio electrónico de transmisión y a la prueba de la reacción en cadena de la polimerasa (PCR). Los resultados mostraron que las muestras reaccionaron fuertemente a antisueros policlonales para la detección de fitoplasmas, se visualizaron cuerpos ovoides y elongados en células de tejido conductor semejantes a fitoplasmas, e iniciadores universales para fitoplasmas en PCR amplificaron un segmento característico de 550 pares de bases. Las evidencias confirman la presencia de fitoplasma en las plantas afectadas. Palabras claves: Vitis vinifera L., reacción en cadena de la polimerasa (PCR) INTRODUCCION El cultivo de la vid (Vitis vinifera L.) es afectado por una serie de enfermedades, cuyos agentes causales pueden ser hongos, bacterias, virus, viroides, fitoplasmas y rickettsias. Los fitoplasmas se encuentran exclusivamente en los vasos floemáticos, normalmente tienen una distribución heterogénea en la planta y se presentan en bajas concentraciones. Estas características hacen difícil su detección e identificación. La asociación de estos organismos con una enfermedad en particular puede estar basada en expresión de síntomas, visualización de estructuras pleomórficas con una membrana trilaminar en las células del tejido conductor del floema, y disminución de síntomas mediante aplicaciones de antibióticos (tetraciclina). En la actualidad, métodos serológicos y moleculares permiten en forma simple, sensitiva y confiable la detección e identificación de estos organismos (Seemuller et al., 1998). Los fitoplasmas causan en las plantas de vides síntomas que incluyen amarillez de hojas, disminución de vigor y bajas de rendimientos; este conjunto de síntomas se denomina "amarillez de la vid" (AV). La primera AV descrita fue la "Flavescence doreé" (FD) en Francia (Caudwell, 1957). Sin embargo, sólo en los últimos años se ha logrado distinguir entre diferentes fitoplasmas causantes de estos síntomas en los cultivos de vides mediante pruebas inmunológicas y otras basadas en ácidos nucleicos. Dentro de las AV se han descrito dos enfermedades muy similares en síntomas, pero que se diferencian en su vector. Una de ellas, FD, es transmitida específicamente por Scaphoideus titanus Ball. (Carraro et al., 1994), mientras que "Bois noir" (BN) y otra enfermedad similar "Vergilbunskrankheit" (VK), son transmitidas por el vector Hyalesthes obsoletus L. (Aleksic et al., 1967). BN fue descrita en 1961 por Caudwell en Francia (Caudwell et al., 1971), mientras que VK fue descrita por Gartel en 1965 en Alemania (Gartel, 1965). Desde el comienzo ambas enfermedades se distinguieron de FD principalmente por su epidemiología, puesto que el vector de FD nunca se encontró presente en vides afectadas con BN o VK. De acuerdo a Boudon-Padieu (1999), las AV también han sido descritas en Rumania (1968), EE.UU. (1977), Australia (1979), Grecia (1979), Italia (1982), Israel (1990), Suiza (1992), Moldavia (1991), Hungría (1997) y Croacia (1991). Por otra parte, Caudwell (1988) y Martelli y Caudwell (1993) indicaron que enfermedades similares a AV están presentes en Chile, Argentina y Australia. Los síntomas de las AV son similares en Norteamérica y Europa, pero el fitoplasma asociado a la enfermedad puede ser diferente. El nombre de FD está restringido a los fitoplasmas de vides transmitidos por S. titanus, y se ha comprobado que pertenecen al grupo de "Elm yellows" (EY). Por otro lado, las AV similares a BN y VK se han asociado a fitoplasmas del grupo de "Stolbur" (Daire et al., 1993). Actualmente, técnicas serológicas y moleculares han proporcionado herramientas para la detección de fitoplasmas, aún en bajas concentraciones, mejorando la seguridad en el diagnóstico. El desarrollo de técnicas tales como la prueba de la reacción en cadena de la polimerasa (PCR), usando iniciadores universales que se aparean en sectores conservados de la región del 16S rADN de procariotes, ha permitido la detección de diferentes fitoplasmas, aún en tejidos donde su concentración es baja (Sinclair et al., 1996). El objetivo de la presente investigación fue identificar mediante métodos inmunológicos, microscópicos y moleculares la presencia de fitoplasmas en plantas de vides. MATERIALES Y MÉTODOS Durante las temporadas 1999 y 2000 se recibieron en el Laboratorio de Virología del Instituto de Investigaciones Agropecuarias (INIA), Centro Regional de Investigación La Platina, Santiago, plantas de vides provenientes de la zona central de Chile, cuyos síntomas correspondían a los descritos para este tipo de patógenos. Las plantas mostraban una declinación progresiva del crecimiento, hojas amarillas y enrolladas hacía abajo, absición prematura de hojas, entrenudos cortos y en general un otoñamiento anticipado. Ante la posibilidad de que el material estuviera infectado con organismos tipo fitoplasma, tanto plantas enfermas como sanas se sometieron a pruebas inmunológicas, moleculares y microscópicas. Inmunología. La prueba inmunológica nitrocelulosa-ELISA (nc-ELISA) se realizó de acuerdo a lo descrito por Conci y Gómez (1999) con algunas modificaciones. Para ello, se maceró 1 g de pecíolos y nervaduras principales de hojas con síntomas en tampón de extracción (1/10 p/v) (0,02 M tris; 0,15 M NaCl; pH 7,5); este macerado se mezcló a razón de 1:1 con cloroformo y se agitó vigorosamente. Enseguida la muestra se centrifugó a 8.000 rpm en una microcentrífuga (Kokusan, modelo H-1500 F, Japón) durante 5 min. Aproximadamente 3 μL de extracto de la muestra y diluciones de 1/20, 1/80, 1/160 y 1/320 en tampón de extracción se colocaron en una membrana de nitrocelulosa, la cual se había sumergido en tampón de extracción durante 1 min. La membrana se dejó secar a temperatura ambiente durante un período de 2 h. Posteriormente la membrana se bloqueó en tampón de bloqueo (0,02 M tris; 0,15 M NaCl; 4% leche descremada y 0,02 g NaN3) durante 2 h. Una vez cumplido este período, la membrana se lavó tres veces con tampón de lavado (0,02 M tris; 0,15 M NaCl; y 0,05% Tween-20) durante 10 min cada vez y se sumergió en inmunoglobulina 1/5000 en una solución conteniendo 2% leche descremada y 0,05% p/v de albúmina de suero de bovino, durante 3 h. Las inmunoglobulinas policlonales utilizadas correspondieron a aquellas producidas en el Instituto de Fitovirología y Fisiología Vegetal (IFFIVE), Instituto Nacional de Tecnología Agropecuarias (INTA), Argentina, para el fitoplasma del paraíso (Conci y Gómez, 1999). Las membranas, una vez lavadas, se sumergieron en una solución de anticuerpo conjugado (anticonejo marcado con fosfatasa alcalina Sigma, diluido 1/40.000 en tampón de inmunoglubulina) durante 2 h a temperatura ambiente. Enseguida, las membranas se lavaron 3 veces durante 10 min con tampón de lavado y se sumergieron en la solución de 10 mg de BCIP/NBT en 10 mL de agua bidestilada estéril (Sigma). Se evaluaron como muestras de reacción positiva todas aquellas que desarrollaron un color púrpura y cuyos controles sanos no mostraron cambios de color. Microscopía electrónica de transmisión. Para la observación ultramicroscópica de cortes ultrafinos se utilizaron pecíolos y nervaduras principales de hojas con síntomas, siguiendo la metodología descrita por Herrera (1990). Para ello, se cortaron trozos de aproximadamente 2 - 4 mm con un bisturí desinfectado. El tejido se fijó con una solución de glutaraldehído al 2% durante toda una noche a 4ºC. Posteriormente, los trozos de tejido se lavaron con tampón fosfato (70 mM NaH2PO4; 30 mM KH2PO2; pH 6,5) tres veces durante 15 min cada vez. Luego, las muestras se sometieron a una post-fijación durante 2,5 h a 4°C, en una solución de tetraoxido de osmio al 1%. El tejido se deshidrató mediante una serie de inmersiones en etanol 50, 70, 90, 95 y 100% de etanol absoluto. Enseguida, se infiltró con resina (Hardener, Ciba, Swiss) a razón de 2:1 (etanol:resina) durante 1 h. Luego, la proporción de etanol:resina se cambió a 1:1 y se dejó durante toda la noche. Finalmente, los trozos se sumergieron en resina al 100% durante 5 h con agitación continua. Cada trozo de tejido, individualmente identificado, se colocó en moldes plásticos, los cuales se incubaron (incubadora IZUSU, modelo 2-2185, Japón) a 70ºC durante 16 h. Los bloques de resina previamente polimerizados en la etapa anterior, fueron tallados en su parte superior en forma piramidal, con el propósito de exponer el trozo de tejido vegetal al cuchillo del ultramicrótomo. Luego, el bloque tallado se colocó en un ultramicrótomo (Leica, modelo S-52, Austria), donde se seccionó el tejido a un grosor de 50 μm. Los cortes desde el micrótomo se colectaron en una grilla (200 mesh) con membrana de Colodium. Los cortes se contrastaron con uranil acetato al 2%, durante 30 min a temperatura ambiente, seguido de un lavado de 1 min en agua destilada y 5 min en citrato de plomo (4 mM Pb(NO3); 6 mM Na3C6H5O7x2H2O; 160 mM NaOH) durante 5 min. Todas las muestras se observaron en un microscopio electrónico de transmisión (Zeiss, modelo EM 900, Alemania). Inmunocaptura seguida de PCR. La aislación de ácidos nucleicos fitoplasmales se realizó mediante el procedimiento de inmunocaptura siguiendo la metodología descrita por Herrera et al. (2001). Para ello, se molieron hojas con síntomas (p/v = 1/4) en tampón de extracción (50 mM citrato trisódico; 1 mM EDTA conteniendo 20 mM dietiltiocarbamato (DIECA) y 2% p/v polivinilpirrolidona (PVP K25, Fluka), el extracto se centrifugó por 10 min a 12.000 rpm. Una alícuota de 100 µL de sobrenadante se colocó en una microplaca, previamente activada con anticuerpo policlonal de fitoplasma (1/5000) proveniente de IFFIVE, INTA, Argentina (Conci y Gómez, 1999) incubándose toda la noche a 4ºC. Posteriormente se lavó con tampón de lavado (0,02 M tris-HCl; 0,15 M NaCl; pH 7,5) tres veces durante 5 min. Enseguida a cada celdilla de la placa se agregaron 10 µL de una solución de Triton X-100 al 10% y se calentó a 65ºC durante 5 min. La amplificación por PCR se realizó colocando 5 μL del producto de la inmunocaptura en una mezcla de 50 μL de reacciσn de PCR (5 μL de tampσn 10X; 2 μL MgCl2 (25 mM); 2 μL de cada iniciador (20 mM); 0,7 μL de dNTPs; 1,0 μL Taq y 28,3 μL de agua estιril). Los iniciadores fueron diseñados de acuerdo a la secuencia del genoma de fitoplasma descrita por Ahrens y Seemuller (1992) (oligo complementario 5´acgaaagcgtggggagaaa´ 3 y oligo homólogo 5´gaagtcgagttgcagacttc´3). La mezcla de PCR se sometió al siguiente esquema térmico; 95°C durante 30 s, 55°C durante 45 s, 72°C durante 30 s (el último de 7 min) y con 35 ciclos. Después de la amplificación los productos de PCR fueron cargados en un gel de poliacrilamida nativo al 5% (38,7 mL de agua estéril; 5 mL de tampón TBE 10X; 6,25 mL de poliacrilamida al 40% (29:1); 50 µL de TEMED (N,N,N',N'tetrametil-etilendiamina) y 340 µL de una solución de persulfato de amonio). El gel se cargó con 8 μL de productos de PCR y 2 μL de tampσn de carga 6X (0,25% azul de bromofenol; 0,25% xilencianol y 30% glicerol en agua) y se corrió a 70 Volts por aproximadamente 2 h en TBE 1X (89 mM Tris-base; 89 mM borato y 2,5 mM Na2EDTA; pH 8,3). La visualización se efectuó por tinción con nitrato de plata; para ello, después de correr el gel, éste se sumergió 1 h en solución de fijación (10% etanol y 1% ácido acético glacial). Luego, se transfirió a una solución de nitrato de plata 0,011 M durante 1 h con agitación constante. Después de lavar dos veces el gel con agua destilada, se agregó el revelador (0,525 M NaOH y 0,002 M NaBH4). El tamaño de los productos fue determinado usando el estándar de peso molecular de ADN 100 bp ladder (Promega, Ma, EE.UU) RESULTADOS Inmunología. Los resultados de las pruebas inmunológicas en base a nitrocelulosa mostraron reacciones positivas para muestras de plantas infectadas con fitoplasmas y reacción negativa para las muestras provenientes de plantas sanas de vides. En las muestras positivas se observó un intenso color púrpura que las diferenció del color café claro en las muestras provenientes de plantas sanas (Figura 1). La afinidad serológica entre las plantas infectadas con fitoplasmas y el antisuero alcanzó hasta diluciones de 1/160 del extracto. Microscopía. Cortes ultrafinos visualizados al microscopio electrónico mostraron cuerpos esféricos, ovoidales y elongados en el citoplasma de células del tejido conductor (Figura 2 A). En algunas células floemáticas estos cuerpos ocupaban toda la cavidad citoplasmática (Figura 2 B). Observaciones más detalladas de estos cuerpos (Figura 2 C y D) mostraron que estaban compuestos de una membrana de característica trilaminar, con dos capas densamente teñidas y otra intermedia de una menor densidad. Frecuentemente los cuerpos ovoides presentaban pigmentación más oscura en su interior, correspondiendo probablemente a ribosomas. El tamaño de los cuerpos ovoidales se presentó variable entre 150 y 400 nm de diámetro, mientras que el tamaño de los elongados llegó a 1500 nm en diámetro. Cortes ultrafinos de células floemáticas de plantas sanas no mostraron ningún tipo de alteración y/o cuerpos semejantes a fitoplasmas. Inmunocaptura seguida de PCR. En la Figura 3 se muestra un gel representativo de los resultados obtenidos cuando el ADN de plantas de vides con síntomas se usó como templado para la acción del par de iniciadores. Consistentemente desde plantas infectadas se amplificó un segmento de ADN característico, de aproximadamente 550 pares de bases (bp). El tamaño de la banda de amplificación correspondió al descrito por Ahrens y Seemuller (1992) cuando el par de iniciadores se enfrenta a ADN proveniente de plantas infectadas con fitoplasmas. En ningún caso se visualizaron productos de amplificación en muestras provenientes de plantas de vides sanas. DISCUSIÓN La presente investigación presenta tres evidencias de la presencia de fitoplasmas en vides. La primera sugiere que plantas con la sintomatología descrita muestran una afinidad con antisueros policlonales (Gomez et al., 1996) para la detección de fitoplasmas. Debido a la presencia, en los fitoplasmas, de una serie de determinates antigénicos comunes, los sistemas de detección serológicos carecen de especificidad para identificar un fitoplasma en particular. Este antisuero generado a partir del fitoplasma que es el agente causal del "declinamiento del paraíso" permite detectar una gama de fitoplasmas que afectan al ajo (Allium sativum L.) y duraznero (Prunus persicaeL.) (Gómez et al., 1996). La segunda evidencia provino de las observaciones en el microscopio electrónico, donde se observó presencia de cuerpos semejantes a fitoplasmas descritos en otros trabajos (Conci y Gómez, 1999) y particularmente en plantas de vides (Parente et al., 1994; Parente y Salema, 2001). La tercera evidencia provino de pruebas de inmunocaptura seguidas de PCR; en esta prueba, la amplificación de segmentos de aproximadamente 550 pares de bases coincidió con los resultados de Ahrens y Seemuller (1992), confirmando la presencia de fitoplasmas en las plantas estudiadas. Sin embargo, la información entregada en la presente investigación no permite concluir respecto del grupo al cual pertenece el o los aislamientos detectados. La literatura reporta a lo menos cinco grupos de fitoplasmas genéticamente distintos, afectando las vides (Seemuller et al., 1998), además, existen evidencias que una misma planta de vid puede ser infectada por fitoplasmas pertenecientes a grupos diferentes (Alma et al., 1996). Los síntomas de FD y BN son similares a los descritos como AV; además es conocido que estas enfermedades son causadas por fitoplasmas. Una amarillez de vid encontrada en Australia (Bonfiglioli et al., 1996), aún cuando se expresó con síntomas similares, su organismo causal, un fitoplasma, era serológica y molecularmente diferente a todos los fitoplasmas conocidos en Europa y EE.UU. (Padovan et al., 1996). La nuevas herramientas moleculares para el estudio de estos patógenos están revelando que existe una gran variedad de fitoplasmas, y que su expresión sintomatológica depende de las condiciones locales en que ellos se expresan (Daire et al., 1997). El principal aporte de esta investigación fue establecer la presencia de fitoplasmas en plantas de vides, y como una consecuencia se plantea la necesidad de realizar los estudios conducentes a caracterizar el o los aislamientos chilenos, estudiar su diseminación, establecer su vector y posibles fuentes reservorios entre las malezas. Las enfermedades causadas por virus y otros organismos afines en el cultivo de la vid son comunes en Europa y EE.UU. (Boudon-Padieu, 1999). La mayoría de los virus más frecuentes en otros países se encuentran presentes en nuestras condiciones (Herrera y Madariaga, 2001), debido probablemente a que al carecer de programas de mejoramiento nacionales, gran parte del material de propagación utilizado tiene su origen en Europa y EE.UU. Es explicable entonces que, al igual que los virus, también se encuentren organismos tipo fitoplasmas afectando las vides. Consecuentemente, los resultados de esta investigación enfatizan la necesidad de promover programas tendientes a la producción de plantas sanas por parte de los viveros. CONCLUSIONES Las evidencias inmunológicas, microscópicas y moleculares confirmaron la presencia de fitoplasmas en plantas de vides afectadas con síntomas de amarillez. LITERATURA CITADA
Full-text article also available online at http://www.inia.cl/at/agritec.htm Copyright 2003 - Instituto de Investigaciones Agropecuarias, INIA (Chile). The following images related to this document are available:Photo images[at03003f2.jpg] [at03003f1.jpg] [at03003f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}