 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
DECOMPOSITION, DYNAMIC OF INORGANIC ELEMENTS AND MICROBIAL COLONIZATION OF ARARIBÁ (Centrolobium tomentosum Guill. ex Benth) LEAVES ON LATOSOLS OF BRAZILIAN ATLANTIC FOREST1Descomposición, dinámica de los elementos inorgánicos y colonización por microrganismos de hojas de araribá (Centrolobium tomentosum Guill. ex Benth) en latosoles del bosque Atlántico brasileño1Márcia S. Marques2, Nadja M. Sá2, and María R. Scotti 2, 3* 1 Recepción
de originales: 06 de agosto de 2001. Code Number: at03008 ABSTRACT Leguminous trees are being planted in order to establish restoration units in the Brazilian Atlantic forest ecosystem. The success of the establishment depends on the growth, productivity of plants and litter decomposition rates. The objective of this study was to investigate the decomposition of araribá (Centrolobium tomentosum Guill. Ex Benth) leaves, at two sites. Site 1 was an experimental plantation of the native species C. tomentosum, which was inoculated with rhizobia plus mycorrhizal fungi, and Site 2 was a eucalyptus (Eucalyptus grandis W. Hill ex Maiden) forest. The leaf litter of C. tomentosum was incubated for 120, 240 and 360 days in both sites. The decomposition rate and dynamic of mineralization of the organic matter and the nutrients K, Mg, Ca dynamics were modified by the site effect. The remaining mass and nutrient content were lower in Site 2 than Site 1. The N and P immobilization occurred eight months after incubation in Site 2 and four months in Site 1. Microbial colonization was made by fungi in Site 2, while in Site 1 the bacteria colonization was higher. The high soil content of Cu, Mn and Zn in Site 2 was not correlated with the metal levels in the residues. The content of Cu and Mn in residues increased in the two sites. Change of eucalyptus plants to trees of C. tomentosum modified the soil biota and favored the bacteria in comparison to fungi. Key words: decomposition, Eucalyptus sp., microbial colonization, tropical forest, soil fertility. RESUMEN Se están cultivando árboles de leguminosas para establecer unidades de restauración en el ecosistema del bosque tropical atlántico de Brasil. El éxito del establecimiento depende del crecimiento, productividad de las plantas, y de la tasa de descomposición de las hojas. El objetivo de este estudio fue investigar la descomposición de las hojas de araribá (Centrolobium tomentosum Guill. ex. Benth), en dos sitios. El Sitio 1 era una plantación experimental de la especie nativa C. tomentosum, inoculado con rhizobios y hongos de la micorriza, y el Sitio 2 era una área con una plantación de eucalipto (Eucalyptus grandis W. Hill ex Maiden). Las hojas caidas de C. tomentosum fueron incubadas durante 120, 240 y 360 días en ambos sitios. La velocidad de descomposición y la dinámica de mineralization de la materia orgánica y de los nutrientes K, Mg y Ca fue modificada por el efecto del sitio. La masa restante y la concentración de nutrientes fueron más bajas en el Sitio 2 que en el Sitio 1. La inmovilización del N y P ocurrió ocho meses después de la incubación en el Sitio 2 y cuatro meses en el Sitio 1. La colonización microbiana fue hecha por hongos en el Sitio 2, mientras que en el Sitio 1 fue por baterias. La alta concentración de Cu, Mn y Zn en el Sitio 2 no tuvo en correlación con los niveles de metales en los residuos. La concentración de Cu y Mn en los residuos aumentaron en los dos sitios. El cambio de plantas de eucalipto a árboles de C. tomentosum modificó la biota del suelo y favoreció a las bacterias en comparación a la población de hongos. Palabras claves: descomposición, Eucaliptus sp., colonización microbiana, bosque tropical, fertilidad de suelo. INTRODUCTION The tropical forest of Rio Doce Park, in the Eastern region of the Brazilian state of Minas Gerais, extends to over 35 974 ha, and it is the largest remaining single block of the Atlantic Forest of Brazil with both primary and secondary vegetation. It is bordered by the Doce River on the East, and Piracicaba River on the North (IEF, 1994) and surrounded by extensive eucalyptus (Eucaliptus grandis W. Hill ex Maiden) plantations which have been planted over the past 30 years. In order to establish eucalyptus plantations, native forests were cut down and the ensuing substitution produced some environmental problems especially in the riparian forests. However, the reforestation companies such as CAF Santa Bárbara (Agroforestry Company Santa Bárbara) and the research institutions, are developing reforestation programmes on the riparian forest of Rio Doce using native species. In some states of Brazil, the preservation of native forest along strips of the rivers and lakes is required by law (Silva Júnior et al., 1995). Multi-purpose trees, particularly those which can fix N are the most interesting.
In 1995, plants of Centrolobium tomentosum Guill. ex Benth, were cultivated
in an experimental area (an abandoned eucalyptus plantation) near River Doce.
C. tomentosum, called "araribá", is a secondary species of Atlantic
Forest, belonging to Papilionoideae, it grows under light, is intolerant to
the
shade (Aidar, 1992), and its primary product is wood for
saw-mill, naval construction and industrial use. The main secondary product is
its high leaf biomass production, which provides larger input of N into the soil.
These effects have been attributed to the double infection of rhizobia and arbuscular
mycorrhizal fungi (AMF). These plants, inoculated with both rhizobia strains BHICBAb1
or BHICBAb3 and AMF, were able to increase their DM production by 56% over the
control plants (Marques et al., 1997, 2001).
The biomass produced depends on the particular species and the nutrient status
of the soil (Jha et al.,1995).
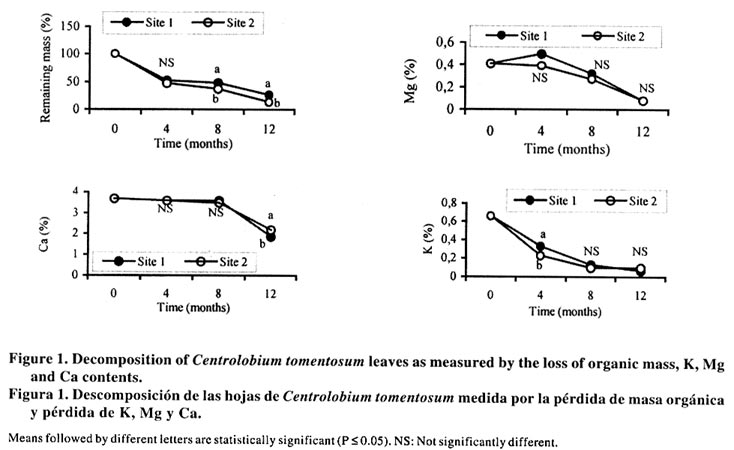
The supply of nutrients depend on organic matter mineralization during its decomposition. Decomposition includes the breakdown of litter and transfers of organic matter and nutrients to the soil. It is an essentially biological process which may be influenced basically by substrate quality and environmental conditions (Swift et al., 1979; Broder and Wagner, 1988). Environmental conditions are related to aeration, moisture condition, pH, and temperature. Substrate quality is related to the C organic chemical composition of the litter. The decomposition rates of leaf litter have been predicted by lignin: N ratio from the leaves (Taylor et al., 1989). In that way leguminous trees that can fix N may increase the nutrient cycling more than non leguminous tree, due to their high N and lignin content as shown by Bernhard-Reversat (1993). On the other hand, changes associated with vegetation cover affect soil organisms and the C decomposition (Tian, 1998). Therefore, variation on land management practices may modify most of the factors which may influence the decomposition process (Mesquita et al., 1998). The primary purpose of this study was to investigate the litter decomposition of C. tomentosum at the site of the experimental plantation and to compare it to the nearest site of eucalyptus forest. MATERIAL AND METHODS Experimental site The study site belongs to CAF Santa Bárbara and is a unit of eucalyptus plantations that has been cultivated since 1970 for charcoal production. This unit was located in the municipality of São José do Goiabal (19°48'-19° 29' S lat; and 42°38''-42°28'W long) in the South East of the Brazilian state of Minas Gerais. The climate is humid-subtropical with a mean temperature for winter of 19-23°C and 25°C for the summer, from 1995 to 1997. Mean annual rainfall is 1450 mm, seasonally distributed, with most rain falling during the summer months (December to February), during the winter exist four to five dry months with a moderate annual water deficit of 30-90 mm. Predominant soil types are Oxisols of various types, and the most frequent of which are the clayey red-yellow latosols (EPFS, 1970). In 1995, an area of 0.67 ha was cleared of eucalyptus plants (Eucalyptus grandis W. Hill ex. Maiden), and was planted with Centrolobium tomentosum Guill. ex Benth, a native leguminous tree of the Atlantic forest, with a density of 1 111 plants ha-1. After the third year of transplanting, the present research was conducted on that area of 0.67 ha, which will be called Site 1. Site 2, located behind Site1, was exclusively composed of eucalyptus trees. Both areas were located 100 m from the river. Soil collection and chemical analysis Soil samples (red-yellow latosol) were collected at a depth of 0-20 cm in Site 1 and Site 2. The soil samples were air dried, sieved (2 mm), and soil analysis were done according to EMBRAPA (1999). Leaf collection Leaves of adult plants of C. tomentosum, which had recently fallen over the ground, were collected in Rio Doce State Park. Leaf litter was collected during August/October 1997. Sample preparation, treatments and experimental design Collected material was air dried and 3 g of leaf samples of C. tomentosum were placed into nylon bags made with 1 mm mesh net. Five sub-samples of each site were retained for initial moisture and chemical analysis. Four bags of each site per each time of analysis (120, 240 and 360 days) were distributed over five randomly located blocks (6 x 4 m) or five replicates, with at least 1 m between each bag. Twenty bags of each site were collected after 4, 8 and 12 months of incubation in the study areas. Sample analysis After being collected the litterbags were transported directly to the laboratory and washed and then brushed in order to remove mineral particles, soil roots and dwarf shrub remnants. About eighteen litter bags per site and time were dried at 850C, individually weighed and then a composite sample was made for each set before the chemical analysis were carried out. Weight loss data was estimated according to Wesemael (1993) using the following equation: % RM= (W0=Wt/X0)*100, where RM = remaining mass; W0 = initial mass; Wt = mass collected al time t. The decay rate (k), which estimates the litter disappearence on a yearly basis and it is proportional to actual decomposition rates, was calculated using the negative exponential decay function done by Olson (1963): k = Wt = W0℮--kt. Litter material was used for analysis of N total nitrogen by Nessler's reagent method (Oliveira, 1986), P by vanado-molybdate method (Sarruge and Haag, 1974), K, Mg, Ca, Zn, Mn and Cu, by nitroperchloric digestion and measured by atomic absorption spectrophotometer (EMBRAPA, 1999). Two litter bags per time and per site were used to estimate the microbial colonization. Microbial population in residues was assessed using the residue washing technique described by Chang and Hudson (1967) on a shaker for 40 min. Procaryotic and eucaryotic populations were determined from solution obtained from the same washings. Serial 10-fold dilutions were conducted with the washings and 1 mL of the appropriate dilutions was used to measure fungi and bacteria (CFU) developed on Martin media (Martin, 1950) and Thorton media (Thorton, 1956) respectively. The cellulolytic population was determined by the most probable number method (Girard-Rougieux, 1964). The analyses of variance were performed using ANOVA (Tukey's multiple-range test). RESULTS AND DISCUSSION The most commonly mentioned factors that may regulate the litter decay are related with litter quality including N elemental concentrations and ratios such as C:N and C:P (Berg et al., 1982; Berg and Ekbohm, 1983; Berg and McClaugherty, 1989); organic matter fractions such as lignin (Meetemeyer, 1978; Taylor et al., 1989), ligno-cellulose index (Berg et al., 1984), lignin:N index (Berg and Ekbohm, 1983; Taylor et al., 1989), alkyl C content of waxes and cutin (Trofymow et al., 1995), elevated CO2 concentration.(de Angelis et al., 2000) and tannin contents (Mesquita et al., 1998). These considerations are more important under litter diversity conditions. However, when the substrate is the same, the chemical composition cannot be correlated to the decomposition rate. In that way it can not explained the differences in decomposition rates of studied sites. On the other hand, the soil type may have influence in decomposition rate. The results (Table 1) showed that there were not many variations in the chemical composition of soil samples of Site 1 and 2, but Site 2, where the eucalyptus forest is, the soil had the highest Zn, Mn and Cu contents. A two-way hierarchical ANOVA showed a significant effect of the site type on the amount of material remaining during 12 months (Table 2). This site effect may be confirmed by the loss of DM of C. tomentosum leaves which is presented in Figure 1. After 12 months the Site 1 residue retained 26% of its original organic matter in comparison with 14 % of Site 2. The more rapid decay in Site 2 started at 8 months and presumably is associated with decomposition of water soluble organic constituents. This hypothesis is reinforced by more rapid loss of K and Mg in contrast with Ca, a constitutive element of middle lamella plants (Figure 1). The more rapid decay in Site 2 may be explained by the annual decay rate (k) of the C. tomentosum leaves in Site 2 (1.82 yr-1) which was higher than Site 1 (1.39 yr-1). These results suggest that soil is a factor which can modify the decomposition rate, but in this case it is not related to the soil chemical properties (Table 1), which were very similar in both Sites. Table 1. Soil chemical parameters
of the studied sites, Minas Gerais, Brazil, 1997.
Table 2. Two-level ANOVA for remaining
mass (%), bacterial, fungi and cellulolytic microbial colonization in leaves
of Centrolobium tomentosum as affected by site and time of incubation,
Minas Gerais, Brazil.
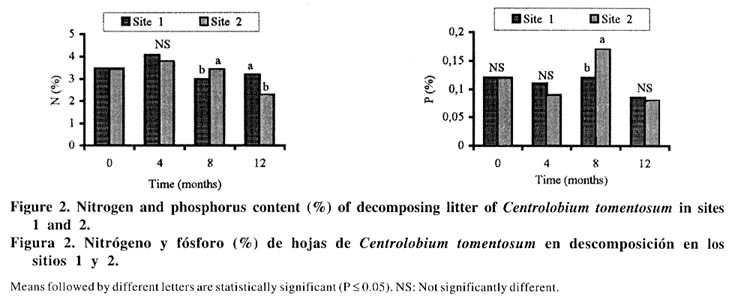
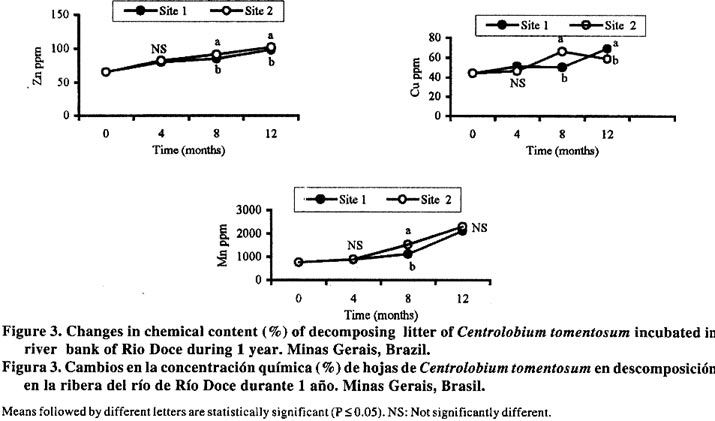
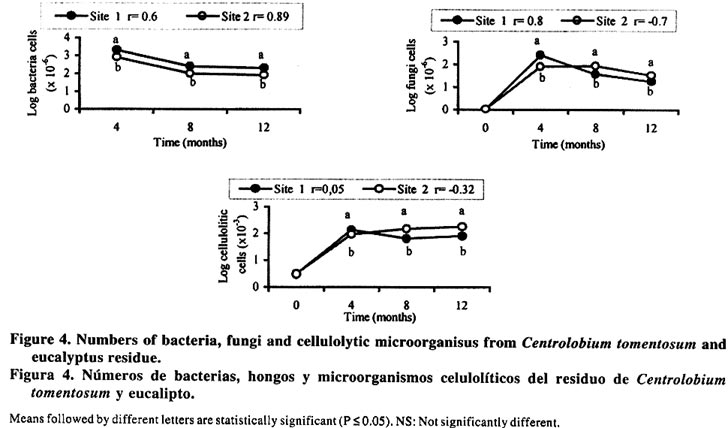
*P< 0.05, **P<0.01, ***P<0.001. The decomposition rate was benefited because the C. tomentosum is a leguminous plant and it has been found that species with a high initial N content as legumes, decompose more rapidly than species with low N (Aber et al., 1990). Nitrogen and P content of litters were not different between the sites during the first eight months of decomposition (Figure 2), but these nutrients were enhanced after eight months of incubation in Site 2. After one year, such increase happened only in Site 1 (Figure 1) and it can be related to nutrient immobilization, specially N. These results suggest that some modification occurred in the C. tomentosum residues after 8 months. In spite of the high metal content in the soil of Site 2, the levels of Cu, Mn and Zn in C. tomentosum residue increased all the time in both soil types (Figure 3). The metal content in litter was not significantly different for Mn, but for Zn it was higher in Site 2 than Site 1. In relation to Cu, their content was enhanced also in Site 2, but only after eight months of incubation. In spite of higher soil content (Table 1), twelve months after the decomposition had started, the pattern changed and Cu content of residues was higher in Site 1 than in Site 2 (Figure 3). Therefore the high metal level in the residues can not be explained by the high metal content in the soil and it reinforces the idea that the soil effect is independent from the nutrients of the soil. Both time and soil type modify the bacteria colonization performance, but no interaction between time and soil was detected (Table 2). Bacteria were the dominant population in decomposing litter of Site 1, and it was independent of the time factor (Figure 4). The high positive correlation (Figure 4) between mass remaining and bacteria colonization in both sites (r = 0.6 and 0.89 for Sites 1 and 2, respectively) showed that bacteria have a more important role in leaves decomposition of C. tomentosum (Table 2). In contrast, time or site may affect the fungi and cellulolityc population, but an interaction between time and site was detected (Table 2). In fact, those populations had been favoured after 8 months of incubation only in Site 2 (Figure 4). The low correlation index (Figure 4) shows that the cellulolytic microorganisms have a little influence in the leaves decomposition of C. tomentosum, contrasting with fungi populations. Fungi colonization decreased during the mass loss (Figure 4) in Site 1 (r = 0.8) while in Site 2 it stayed high (r = -0.7). These results can suggest that succession of microorganism population was different for each site. Organic matter decomposition is considered a functional indicator of ecosystems (Odum, 1983). Variation in land management practices changes most of the factors that influence residues decomposition. Leaf decomposition and nutrient release are lower in a degraded soil than in a non-degraded soil, and it has been attributed to the reduction of soil biota (Tian, 1998). Therefore, it was expected that soil microorganisms may present differences between Eucalyptus sp. and C. tomentosum sites. These results showed that bacteria had a greater influence in Site 1 with time, while in Site 2 fungi were more important decomposers after eighth month, mainly during the rainy season. The remainder leaves of the (undecomposed) in Site 2 disappeared more rapidly than in Site 1 after eighth month. This increase in the decomposition rate coincided with an increase in microbe colonization after four months for Site 1 and after eighth months of incubation for Site 2. These data shows that fungal-based colonization was responsible for this high degree of immobilization and it may be related with the higher mass of filamentous body as observed by Paustian (1985). After twelve months of incubation, the number of fungi stayed higher in Site 2 than in Site 1, but bacteria and fungi populations were reduced in relation to the previous measurement. The high number of cellulolytic cells imply a persistent activity over cellulose residues specially in Site 2, where the residue are richer in cellulose. However, the amount of mass remaining and N content of residues were greater in Site 1 than Site 2, so it can not be attributed to microorganisms immobilization, but it may be probably related with the formation of refractory component which can retain N. The fact that the same litter was colonized differently in the two sites, suggests that the resident microbial populations would be different and the final product of decomposition may also be different. The number of bacteria cells on decomposing litter on Site 2 was lower than Site 1. These results suggest that a selective pressure occurred in Site 2 and it could have favoured the fungi population as compared to the bacteria and cellulolytic microorganisms. It is known that eucalyptus produces inhibitory substances for the bacteria growth, and under this condition the fungi population may be favoured (Della-Bruna et al., 1989; Moura et al., 1996). Furthermore, during the experiment, the content of Mn and Zn increased especially where the eucalyptus trees were planted. This increase was found also by Muoghalu et al. (1994), whom observed that Mn, Cu and Zn increased in concentration during the litter decomposition. According to Muoghalu et al. (1994) this increase may be also related to the retention as complex forms with the organic constituent. Besides, Prosser et al. (1993) and Muniz et al., (1996) observed that in eucalyptus soils the levels of Al, Mn and Zn associated with the organic matter were larger than the average of other soils due the formation of stable complexes between organic matter and transition metals. These results indicate that the residual organic matter of Site 2 would be capable of retaining more Zn while in Site 1 more Cu is complexed. The results presented here suggest that a high rate of leaf decomposition in Site 2 was caused by fungi population, which was improved by selective pressure in eucalyptus site. Restoration techniques using native species as C. tomentosum, which showed a low C:N relationship, may be able to modify the decomposer communities and decomposition rates. CONCLUSION
ACKNOWLEDGEMENTS
This study was supported by Brazilian Conselho Nacional de Pesquisa (CNPq), Fundação de Amparo a Pesquisa do Estado de Minas Gerais (FAPEMIG) and Pró-Floresta Program of World Bank and Instituto Estadual de Florestas grant and to CAF Santa Bárbara Company. LITERATURE CITED
Full-text article also available online at http://www.inia.cl/at/agritec.htm Copyright 2003 - Instituto de Investigaciones Agropecuarias, INIA (Chile). The following images related to this document are available:Photo images[at03008f3.jpg] [at03008f1.jpg] [at03008f4.jpg] [at03008f2.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}