 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
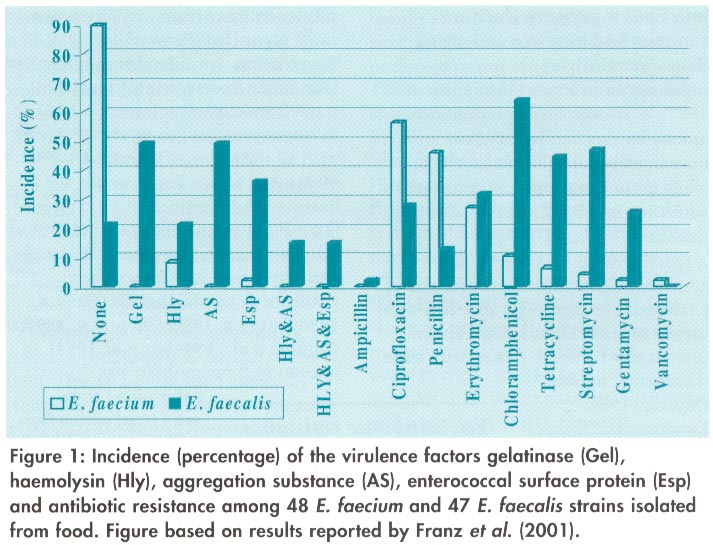
Australasian Biotechnology, Vol. 12 No. 1, 2002, pp. 31-37 BIOTECHNOLOGY IN FOOD PRODUCTION OF ENTEROCOCCI AND FOOD Charles M.A.P. Franz, Hikmate Abriouel and Wilhelm H. Holzapfel Federal Research Centre for Nutrition, Institute ofBiotechnology and Molecular Biology, Haid-und-Neu Stasse 9, D-76131 Karlsruhe, Germany, and Antonio Gdlvez, Departmcnto Microbiologia, Universidad deJaén, 23071 Jaén, Spain, Email: Wilhelm.Holzapfrl@bfe.uni-karlsruhe.de Code Number: au02008 Enterococci are lactic acid bacteria of importance in foods; they can spoil processed meats but are also important for the ripening and aroma development of certain Mediterranean cheeses and sausages. Enterococci have also been successfully used as human probiotics. However, they are also recognised as important nosocomial pathogens causing bacteraemia, endocarditis and other infections. Some strains are multiply antibiotic resistant, but antibiotic resistance alone cannot explain the virulence of some strains of these bacteria. Virulence factors such as production of adhesins, invasins, and hemolysin have now been described. The role of enterococci in disease has raised questions on their safety for use in foods or as probiotics. Recent studies on the incidence of virulence traits among food strains showed that some food isolates can also harbour such traits. Generally, E. faecalis appears to harbour more virulence traits than E. faecium and they occur at a greater incidence among E. faecalis strains. Regulation in Europe stipulates that safety of probiotic or starter strains is the responsibility of the producer; therefore, each strain intended for such use should be carefully evaluated. The Enterococci Enterococci are typical lactic acid bacteria (LAB) and are of importance in food and clinical microbiology. Representatives of the genus Streptococcus formerly grouped as ‘faecal streptococci’ or Lancefield’s group D streptococci, were separated from this genus on the basis of modern classification techniques and serological studies in the 1980’s. The large conglomeration of streptococci was thus subdivided into three separate genera: Streptococcus, Lactococcus and Enterococcus (Schleifer and Kilpper-Balz, 1984; Devriese et al, 1993; Devriese and Pot, 1995). The typical pathogenic pathogenic species, with the exception of S. thermophilus, remained in the genus Streptococcus, and were separated from non pathogenic and technically important species of the new genus Lactococcus (Devriese and Pot, 1995). The 'faecal streptococci' associated with the gastrointestinal tract of man and animals, with some fermented foods and a range of other habitats, constitute the new genus Enterococcus. Although more than 20 Enterococcus species are currently recognised, E. faecium and E. faecalis are still the two most prominent representatives and are the species which play the most important roles both in human disease and in fermented foods and in probiotics (Franz et al. 1999a). Beneficial activities of Enterococci Enterococci in food production. Enterococci are associated with traditional European cheeses produced in Mediterranean countries from raw or pasteurised ewe’s or goat’s milk. The enterococci involved either originate from environmental contamination or from raw milk and may well survive mild heat treatments (Cogan et al. 1997; Garcia-Armesto et al, 1993). Enterococci (especially E. faecium and E. faecalis) are part of the natural microbiota of the ruminal ecosystem both in wild and domestic ruminants (Lauková, 1996), and they are released in large numbers in faeces, from where they may contaminate the udder and the milk. Enterococci are able to grow in the rather restrictive environment of high salt content and low pH of such cheeses (Ordoflez et al. 1978; LitopoulouTzanetaki, 1990, Wessels et al. 1990, Freitas et al. 1995). Most importantly, the enterococci contribute to the ripening and aroma development of these products (Jensen et al. 1975; Ordoflez et al, 1978; Trovatelli and Schiesser, 1987; DeFernando et al. 1992; Centeno et al. 1996), so that organoleptically unique products are produced which contribute to the local cuisine and heritage of the region. in addition, due to interlinked European and worldwide markets, these cheeses are widely distributed and are internationally considered as delicacies. One special benefit of the presence of enterococci in cheeses, is that such strains also may have bactertocin activity. Bacteriocins are antimicrobial peptides (small proteins) which can inhibit the growth of related strains, including some food pathogenic bacteria (Klaenhammer, 1993). Because of this antimicrobial activity towards some foodborne pathogens, bacteriocins are often considered as natural or ‘bin’ preservatives. Adulteration of cheeses by L. monocytogenes can also occur and contaminated cheeses have in the past led to disease outbreaks. Therefore, research has focused on the use of naturally occurring, bacteriocin-producing enterococci as starter cultures for such food commodities, in order to combat the growth and dissemination of this pathogen in particular. Bacteriocin production by enterococci isolated from dairy products has been investigated in detail for a number of strains, and these were shown to produce a variety of different bacteriocins. Strains producing the broad-spectrum, plasmid-encoded cyclic bacteriocin AS-48 (Gálvez et al, 1986; Gonzalez et al. 2000; MartinezBueno et al. 1998) have been found in raw milk and dairy products (MaisnierFarm et al, 1996; Maqueda et al, 1998; Rodriguez et al, 1998; Rodriguez et al, 2000). AS-48 shows a broad spectrum of antimicrobial activity (Gálvez et al, 1996), even towards foodborne pathogens such as L. monocytogenes (Garcia et al, 1997; Mendoza et al. 1999), B. cereus (Abriouel et al, submitted), and permeabilised cells of Salmonella choleraesuis (Abriouel et al, 1998). The use of starter cultures containing the bacteriocin-producing enterococci for cheese making to improve their safety has been described (Sulzer et al. 1992; Garcia et al. 1997; Oumer et al. 2000). Enterococci have been isolated also from Spanish-style green-olive fermentations (Fernández Diez, 1983; Floriano et al. 1998), in which E. faecalis is a frequent contaminant, and they frequently occur in retail fermented olives (unpublished results). However, the role of enterococci in olive fermentation is largely unknown. it has been suggested that enterococci can use the antimicrobial glucoside oleuropein found in olive grapes as a growth substrate (Garrido-Fernandez and Vaughn, 1978), lowering the toxicity of the fermentation medium for other LAB. Bacteriocinproducing enterococci have also been described for other vegetable products (Floriano et al. 1998; Franz et al. 1996; McKay, 1990). Fnterocin B produced by F. faecium BFE 900 isolated from black olives (Franz et al. 1999b) and enterocin I produced by E. faecium 6T1 from a Spanish-style green-olive fermentation (Floriano et al. 1998) have been studied in detail. Enterocin I is active against many olive spoilage and food-borne pathogenic bacteria, including clostridia, propionibacteria, Bacillus, and Listeria monocytogenes (Floriano et al. 1998). Enterococci are also isolated from certain types of fermented sausages, particularly the dry fermented sausages known as ‘chorizo’ (Casaus etal, 1997; Cintas et al. 1997) and ‘espetec (Aymerich et al. 1996) produced in Spain. Production of ant i-listerial bacteriocins by various enterococci from Spanish-style dry fermented sausages suggests that such strains may be suitable for addition to meat as cocultures to improve food safety (Casaus et al. 1997; Callewaert et al. 2000; Cintas et al. 2000). Enterocins A and B from E. faecium CTC492 (Aymerich et al. 1996) show marked antilisterial activity in meat and meat products such as cooked ham, minced pork meat, deboned chicken breasts, pate, ‘espetec’, and dry fermented sausages, and it has been proposed that they could be considered as additional biopreservative hurdles for prevention of listerial growth (Aymerich et al. 2000 a and b). Enterococci as probiotics By definition, probiotics are ‘a mono or mixed culture of live microorganisms which, when applied to animal or man, beneficially affect the host by improving the properties of the indigenous flora’ (Havenaar et al. 1992). Suggested functional effects contributed by probiotics include: inhibition of pathogenic microorganisms, strengthening of the gut mucosal barrier, antimutagenic and anticarcinogenic activities, stimulation of the immune system and lowering of blood cholesterol levels (Fuller, 1989 Havenaar and Huis in ‘t Veld, 1992; Pool-Zobel et al. , 1993; Salminen et al. 1993, 1996; Holzapfel et al. 1998). Most probiotic strains are of intestinal origin and belong to the genera Bifidobacterium and Lactobacillus. Enterococci, as natural inhabitants of the human gastrointestinal system, are also used occasionally as probiotics. One of the best-studied enterococci used as a probiotic is E. faecium strain SF68. This strain has been especially used for the treatment of diarrhoea and is considered as an alternative to antibiotic treatment for this affliction (Lewenstein et al. , 1979; Bellomo et al. 1980). Controlled ‘double blind’-type clinical studies have shown that treatment of enteritis with E. faecium SF68 was successful in both adults and children and decreased the duration of diarrhoea and the time for normalisation of patient’s stools (Bellomo et al. , 1980, Bruno and Frigerio, 1981; D’Apuzzo and Salzberg, 1982). The use of enterococci as probiotics is currently a controversial issue. On the one hand, the beneficial aspects for some strains used in probiotic preparations has been proven and, for example, the SF68 strain has a long history of safe use. On the other hand, the emergence of multiply antibiotic-resistant enterococci in recent years and the increased association of enterococci with human disease (see below), has raised concern regarding the use, especially of ‘new strains, as probiotics. Detrimental activities of enterococci Enterococcal Food contamination and Food spoilage Enterococci constitute a large proportion of the autochthonous bacteria associated with the mammalian gastrointestinal tract (Devriese et al ., 1991, 1992, 1994; Leclerc et al. , 1996). Thus, the presence of enterococci in the gastrointestinal tract of animals clearly leads to a high potential for contamination at the time of slaughter. In raw meat products, F. faecalis was shown to be the predominant isolate from beef and pork cuts in one study (Stiles et al. 1978), while in another both E. faecium and E. faecalis were the most predominant Enterococcus spp. isolated from pig carcasses (Knudtson and Hartman, 1993). These pig carcasses from three different slaughter plants contained mean counts of 104 to 108 enterococci per 100 cm2 of carcass surface throughout processing (Knudtson and Hartman, 1993). In a study on poultry, E. faecalis predominated among the Gram-positive cocci isolated from chicken samples collected at abattoirs (Turtura and Lorenzelli, 1994). However, enterococci may not only contaminate raw meats, but they can also be associated with processed meats. Cooking of processed meats may confer a selective advantage on enterococci as these bacteria are known to be among the most thermotolerant of the nonsporulating bacteria (Sanz Perez et al. 1982, Magnus et al. ., 1988). After surviving the heat-processing step, both E. faecalis and E. faecium have been implicated in spoilage of cured meat products such as canned hams and chub-packed luncheon meats (Bell and Gill, 1982; Houben, 1982; Bell and Delacey, 1984; Magnus et al. 1986). Effective control of meat contamination by enterococci may become more important in future with increased recognition of these bacteria as human opportunistic pathogens and dissemination of antibiotic resistance genes (see below). Enterococci as opportunistic pathogens of humans In recent years, enterococci have become recognised as major nosocomial pathogens causing bacteraemia, endocarditis as well as urinary tract and other infections (Murray, 1990; Morrison et al. 1997). In the USA, their importance has increased from the third leading cause of nosocomial disease in 1984 to the second between 1985 and 1989 (Chenoweth and Schaberg, 1990; Schaberg et al. , 1991). Enterococci are opportunistic pathogens and cause infections in patients which are compromised by severe underlying disease or which are immunocompromised (Morrison et al, 1997). E. faecalis clearly predominates among enterococci isolated from human infections (more than 80%), while E. faecium strains are associated with the majority of the remaining infections (Jett et al. 1994). A specific cause for concern and contributing factor to pathogenesis of enterococci is their resistance to a wide variety of antibiotics, which renders infections with some strains with little therapeutic options (Murray, 1990; Landman and Quale, 1997; Leclercq, 1997). It is well known that enterococci are naturally resistant towards a number of antibiotics. It is, therefore, important to distinguish between this type of intrinsic resistance which is generally mediated by genes located on the chromosome, and acquired resistance, which is mediated by genes residing on plasmids or transposons (Clewell, 1990; Murray, 1990). Examples of intrinsic antibiotic resistance to cephalosporins, b-lactams, sulphonamides and low levels of clindamycin and aminoglycosides, while examples for acquired resistance include resistance to chloramphenicol, erythromycin, high levels of clindamycin and animoglycosides, tetracycline, high levels of b-lactams, fluorquinolones and glycopeptides such as vancomycin (Murray, 1990; Leclercq, 1997). Vancomycin resistance is of special concern as this was considered as a last resort for treatment of multiple resistant enterococcal infections, and the emergence of vancomycin resistant enterocci (VRE) in hospitals has led to infectinos untreatable with antibiotics. Yet, antibiotic resistance alone can not explain the virulence of enterococci. The pathogenesis of most infections follows a common sequence of events involving colonisation of host tissues, invasion of the tissue and resistance to both non-specific and specific defense mechanisms which are mobilised by the host. Finally, the pathogen must be able to produce pathological changes either directly, by producing toxin, or indirectly by producing inflammation (Johnson, 1994). Enterococci possess subtle virulence traits, which are not easily identified (Jett et al. 1994). Nevertheless, considerable progress has been made in determining virulence traits from clinical enterococci isolates in recent years and these are associated with all the stages of infection mentioned above. For example, the bhaemolysin-bacteriocin (Cyl, cytolysin) is a cellular toxin produced by a number of clinical isolates of E. faecalis and was shown to enhance their virulence in animal models (Ike et al., 1984; Jett et al, 1992; Chow et al, 1993; Gilmore et al. 1994; Jett et al, 1994). In one study, bhaemolysin was determined to beassociated with a fivefold increased risk of death within 3 weeks of bacteraemia compared with patients with bacteraemia caused by non-b-haemolysin producing strains (Huycke et al. 1991). Aggregation substance (AS) is an adhesin that is encoded on pheromone responsive plasmids. It mediates the formation of cell clumps, which allows the highly efficient transfer of the sex pheromone plasmid on which it is encoded (Clewell, 1993). AS was shown to facilitate adherence to and invasion of eukaryotic cells in tissue culture experiments (Kreft et al. 1992; Olmsted et al. 1994) and, therefore, is an important virulence factor as it facilitates both adherence (adhesin) to and invasion (invasin) of cells. Gelatinase is a proteolytic enzyme (extracellular metalloendopeptidase) that acts on collagenous material present in tissues. The production of gelatinase was shown to increase pathogenicity in an animal model (Singh et al. 1998) and this confirmed its role in virulence of enterococci. Although production of gelatinase, b-haemolysin and AS are well-defined virulence traits, the role of which in pathogenicity has been confirmed, other virulence factors are less well studied. One such potential virulence factor is an adhesin, the ‘enterococcal surface protein’ (Esp), which is encoded on the chromosome. The incidence of Esp was shown to be higher among clinical E. faecalis strains when compared to E. faecalis strains isolated from healthy individuals (Shankar et al. 1999), indicating that this potential virulence trait is important in disease. The Esp protein consists of a core region that consists of repeat units. The repeat units suggest that different Esp+ enterococci may exhibit variations in these repeats, which may lead to the expression of variant proteins which are identical at the amino and carboxy termini, but differ in the number of repeats (Shankar et al. 1999). Such a phenomenon is thought to be related to evasion of the immune response (Shankar et al. 1999). Other potential virulence factors, the role of which has not yet been established in animal models, include the production of a collagen binding protein termed 'adhesin of collagen from E. faecalis’ or Ace (Rich et al. 1999). Recently, it was shown that Ace not only binds to the extracellular matrix proteins collagen (types I and IV), but also to laminin (Nallapareddy et al, 2000 a,b). Nallapareddy et al (2000b) also showed that Ace was expressed during infections, as human sera collected from patients with enterococcal infections reacted with anti-Ace antibodies. Production of the adhesin-like E. faecalis and E. faecium endocarditis antigens (EfaAtfs and EfaAfm, respectively) are also considered as potential virulence determinants. EfaAfs was suggested to play a role in adhesion in endocarditis (Lowe et al. 1995) and, so far, only the efaAfs, gene was shown to influence pathogenicity in animal models (Singh et al. 1998). Incidence of virulence factors among food enterococci strains Considerable progress has been made in the last few years in determining virulence factors from clinical enterococci isolates using molecular biological techniques and model animal experiments. As a result, we now have the opportunity for studying the incidence of such virulence factors among food isolates and determine whether there is a difference in virulence potential between food and medical strains. In addition, such studies may allow evaluations on whether a strain that is intended for use as a probiotic or starter culture would be safe. One such study compared the incidence of virulence factors among enterococci strains intentionally added to foods as starter strains, as well as food and medical isolates (baton and Gasson, 2001). This study showed that enterococcal virulence factors occurred among strains from all three sources, but that the incidence of virulence factors was highest for medical strains, followed by food isolates and the lowest incidence was observed for starter strains. E. faecalis isolates harboured multiple virulence determinants while E. faecium strains, with the exception of a few strains, were generally clear of virulence determinants (Eaton and Gasson, 2001). A similar low incidence of virulence factors among E. faecium strains isolated from food was observed in our studies, with only few strains producing either hemolysin (8.3%) or Esp (2.1%) (Franz et al. 2001). However, E. faecalis strains in our study also harboured multiple virulence determinants, and the incidence of these was much higher than for E. faecium (Figure. 1). This could be explained on the basis that some of these virulence determinants (e.g. aggregation substance, cytolysin) are encoded on pheromone-responsive plasmids, which facilitate transfer of plasmid DNA at a high frequency. These pheromone-responsive plasmids, however, generally occur only for E. faecalis, although the presence of such a plasmid transfer system was described for E. faecium also in a single report (Heaton et al. 1996). In our studies we also investigated the incidence of antibiotic resistance among enterococci, and found that many strains showed resistance to one or more of the antibiotics (Fig. 1). Still, the majority of the food enterococci isolates, especially the E. faecium strains, appeared to be sensitive to the clinically relevant antibiotics ampicillin, streptomycin, gentamycin and vancomycin (Fig. 1). Safety of enterococci strains for use in foods or as probiotics These first investigations suggest the incidences of virulence determinants among food isolates to be strain-specific (Franz et al, 20001). For this reason, should an Enterococcus be considered for use as a starter culture or as a probiotic, each strain would have to be carefully evaluated for its virulence potential. In general, E. faecium strains appear to pose a lower risk for use in foods, as these strains generally harbour less virulence determinants than E. faecalis and the incidence of virulence determinants among these strains is low. The opposite applies to E. faecalis strains, and this may be a result of the presence of pheromone-response plasmids, which may encode virulence factors. As enterococci possess different gene transfer mechanisms (e.g.. pheromone-responsive plasmids, conjugative and non-conjugative plasmids and transposons) it is feared that enterococci lacking virulence determinants may acquire these from other enterococcal strains. This would clearly be one of the risk factors for the use of enterococci that do not display any virulence determinants as probiotics or starter cultures. Again, such a risk would be higher fur E. faecalis strains, as the gene transfer mechanism based on pheromone response plasmids is known to generally occur in this species (Clewell, 1993). Indeed, Eaton and Gasson (2001) highlighted this probability of gene transfer by showing that virulence genes on a pheromone response plasmid could be transferred to food E. faecalis starter strains. However, they were unable to transfer virulence genes into a food E. faecium starter strain (Eaton and Gasson, 2001). Regulatory aspects Presently, only the use and application of animal probiotics are regulated in Europe. This is based on Council directive 70/524/EEC of 23rd November 1970 on feed additives and which was recently amended by directive 1999/20/EC. Regulation and formal mandatory approval of new LAB starter cultures for human use do not exist in the same way as it is applied to animal probiotics. This practice is based on the long tradition of safe use and association of LAB in food fermentations and their ‘GRAS’ status. This also applies to new probiotic strains, although, in terms of consumer protection, proof of both safety and functionality is considered to be the responsibility of the manufacturer. Present regulations regarding labelling and the prohibition of health claims are considered to provide sufficient consumer protection in terms of the EU guideline 79/112/EEC. Clearance of new strains for application as either starter cultures or probiotic cultures, may be sought for example from the British Advisory Committee on Novel Foods and Processes (ACNFP). An example is E. faecium strain K77D for use as a starter culture for fermented milk products. After clearance was obtained for its use in an EU member state (Denmark), according to EU legislation, marketing of the product is legal in all other member states. The ACNFP evaluation focused particularly on the strain’s history, its identification (by I)NA fingerprinting) as well as specification and quality control criteria. Evidence was also provided to demonstrate the lack of pathogenicity and the lack of any adverse effects in volunteers during a six-week trial. As the onus is on producers of starter cultures and probiotics to show that their strains are safe for use, rigorous testing for all known virulence traits and antibiotic resistances for each strain is required before such a strain is employed. Absolute safety, however, can never be assumed, as undetected virulence traits may exist and transfer of virulence determinants could theoretically occur. These considerations are of particular importance with regard to the risk groups in the population (the young, old, those with underlying disease and mmmunocompromised) but may not necessarily have consequences for the healthy population. Acknowledgement Our study on the incidence of virulence factors among enterococci isolated from food, was carried out with the finammcial support from the Commission ofthe European Communities, Agriculture and Fisheries (FAIR) specific RTD programme under the project CT97-3078 Enterococci in Food Fermentations. Functional and Safety Aspects. It does not necessarily reflect its views and in no way anticipates the Commission’s future policy in this area. References
Copyright 2002 - AusBiotech The following images related to this document are available:Photo images[au02008f1.jpg] |
| |||||||||
{kind=link}