 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Volume 6 Number 3, May/June 1996,pp.168-173 Glycobiology: the technologies associated with glycoprotein analysis Andrew A. Gooley, Anthony Pisano, Nicolle H. Packer, John W. Redmond, Warren Kett and Keith L. Williams Macquarie University Centre for Analytical Biotechnology, School of Biological Sciences, Macquarie University, Sydney, NSW, 2109
Code Number: AU96003
Size of Files:
Text: 20.9K
Graphics: Line drawimgs (gif) - 52.3K
[ALL TABLES AND FIGURES AT END OF TEXT] Protein glycosylation is ubiquitous being found on molecules in both eukaryotes and to an increasing extent prokaryotes. Why are proteins modified with sugars? One general consequence for glycosylation on proteins is that the glycosylation imparts proteinase resistance. There are numerous examples of glycoprotein instability following the site directed metagenesis of a glycosylation site on proteins such as the GLYT1 glucose transporter protein, Human transferrin receptor and the angiotensin-converting enzyme, all expressed in chinese hamster ovary cells. However, increasingly glycosylation on proteins has been found to be important for protein trafficking, biological activity, cell-cell and cell-extracellular matrix interactions (Varki, 1993). The broad interest in the structure and function of glycoproteins has led to a relatively new discipline, glycobiology. The principal aim of glycobiology is to understand the structure and function of the carbohydrate attached to a variety of carriers including protein (glycoproteins, proteoglycans), lipid (lipopolysaccharides, glycolipids) and to protein and lipid (glycophosphatidyl inositol protein anchors, lipopeptidophosphoglycans). Understanding glycosylation patterns is also of relevance to recombinant protein design where it is desirable to engineer glycoproteins with a variety of physio-chemical properties including proteolytic stability, thermostability, elasto-viscosity and structural modulation of receptors (Dwek et al. 1993). Glycobiology is an emerging field and its development remains dependent on improving the accuracy and sensitivity of carbohydrate analysis. The traditional approach to the compositional and structural characterisation of glycoproteins has been to treat both the polypeptide chain and the carbohydrate component as separate entities. This paper reviews the current status of the technologies associated with glycoprotein analysis and an exciting new approach is considered where the glycoprotein is analysed with the carbohydrate still attached. This provides a level of detail once thought not possible.
Methods for the characterisation of carbohydrate attached to glycoproteins
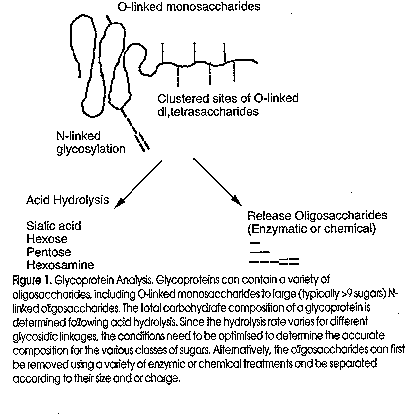
Figure 1 shows a cartoon of the current approach to the analysis of the sugar component of a glycoprotein.
Monosaccharide analysis Monosaccharides, released by acid hydrolysis, have historically been separated on paper or thin layer chromatography using colour reactions to detect different classes of sugars. Advances in high performance liquid chromatography (HPLC) technology have led to the development of monosaccharide separation methods based on a variety of different columns and a range of derivatisation and detection techniques. These have included partitioning in both normal and reversed phase, anion and cation exchange, ligand exchange and, most recently, high pH pellicular anion exchange. The major problem in oligosaccharide compositional analysis is that glycosidic bonds hydrolyse at highly variable rates. The liberated monosaccharides also can decompose under prolonged or severe hydrolysis conditions. In any hydrolysis there is a tradeoff between incomplete cleavage and decomposition of the components. Sialyl and furanose bonds are the most acid labile and glucosaminyl and uronic acid bonds are the most acid resistant, with uronic acid then becoming susceptible to decarboxylation once released. If the amino sugars are acetylated there can be a loss of the acetyl group during the hydrolysis, which renders the glucosaminyl bond more acid resistant.
The most often used acid catalysts are hydrochloric and trifluoroacetic acid. Due to its volatility and consequent ease of removal the most commonly used hydrolysis conditions involve 2 M trifluoroacetic acid at 100 C for 4 hours, which is reported to give quantitative recovery of neutral monosaccharides and about 75% recovery of aminohexose residues (depending on linkage) (Hardy et al, 1988). Under such conditions sialic acids are completely destroyed, but these can be analysed using milder acidic conditions for example, 0.1 M trifluoroacetic acid at 80 C for 45 minutes.
Release of Carbohydrate from Glycoproteins
Before the structure of an oligosaccharide can be determined the sugar chain must be released from the molecule to which it is attached. Each glycoconjugate has unique characteristics which make it impossible to give a general method of release of the glycan moiety. Carbohydrate covalently linked to glycoproteins can be divided into three major classes:
ii) O-linked glycosylation (or O-glycosylation) is where a monosaccharide is covalently attached to the hydroxyl side-chain of an acceptor Hydroxyamino acid residue, usually a Serine or Threonine and in some cases a Hydroxylysine and Hydroxyproline followed by (in many cases) stepwise addition of further sugar residues.
iii) glycosylphosphatidylinositol (GPI) anchor, where a dolichol precursor oligosaccharide is covalently attached via an ethanolamine phosphate to the C-terminus of the protein. To determine the complete structure of carbohydrates attached to glycoproteins, it is necessary to first release them from the peptide backbone and then determine which residues are present, which linkages are present, and whether each amino acid attachment site contains a homogeneous population of carbohydrate. There are a variety of enzymatic and chemical methods used for the release of oligosaccharides and often several methods are needed for the complete characterisation of a proteins carbohydrate complement (Figure 2). For the release and recovery of most N-glycans, the preferred method is the enzymatic release with PNGase F, which hydrolyses the b-aspartylglucosamine bond between GlcNAc and Asn. In contrast, the only available O-glycan specific endoglycosidase (Endo-a-N-acetylgalactosaminidase) is highly specific and only cleaves mucin-type Galb1-3GalNAc-Ser/Thr linkages. It does not cleave if the disaccharide is modified in anyway, for example by sialylation, sulfation or elongation. As there are at least eight different reducing terminal sugar linkages to Serine and Threonine (Gooley and Williams, 1994), chemical release of O-glycans is more commonly used.
Oligosaccharides are released by hydrazinolysis (N & O-glycans) as glycan hydrazones and recovered as reducing sugars (Patel and Parekh, 1994). The conditions for release can be optimised so that there is selective removal of O-glycans from glycoproteins which also carry N-glycans. The chemical cleavage requires high grade hydrazine with salt-free lyophilized glycoprotein/glycopeptide and usually removes 90% of N-/O-glycans. Alkaline borohydride b-elimination recovers O-linked oligosaccharides as sugar alditols.
Methods for the Characterisation of Released Glycans from Glycoproteins
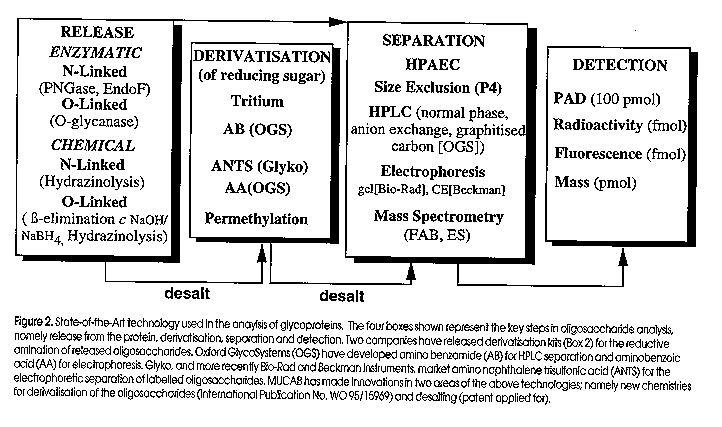
Oligosaccharides recovered from the enzymatic or chemical treatment of a glycoprotein are separated by liquid chromatography or electrophoresis and then detected using a variety of methods (Figure 2). Except for pulsed amperometric detection (PAD), all oligosaccharides and monosaccharides must first be chemically derivatised at the free reducing sugar prior to separation. The derivatisation chemistries include reductive amination (amino benzamide, AB; amino naphthalene trisulfonic acid, ANTS; amino benzoic acid, AA), permethylation (NaOH/methyl iodide), or sodium borotritide (NaB3H4) reduction of the alkaline beta-eliminated carbohydrate. All of these derivatisation procedures allow detection in the pmol to fmol range.
One of the most popular methods, albeit less sensitive, for the analysis and detection of both oligosaccharides and monosaccharides is high performance anion exchange chromatography on a high pH stable pellicular resisn, with pulsed amperometric detection (HPAEC-PAD). The advantage of the technique is its speed, high resolution separation of both monosaccharides and oligosaccharides, and that there is no need to derivatise the sugars prior to separation and detection. The method is also compatible with glycoproteins separated by gel electrophoresis and electroblotted onto polyvinylidene difluoride (PVDF, Packer et al, 1996).
One of the most promising separation new methods for carbohydrates is electrophoresis. The popularity of polyacrylamide gel electrophoresis (PAGE) based separations and the high resolution capacity of capillary zone electrophoresis (CZE) may provide the incentive for methods development so that carbohydrate analysis becomes routine rather than a specialist technique centralised in core facilities. Several manufacturers now market kits for labelling carbohydrates with a fluororphore (eg: ANTS and AA, see Figure 2) for separating the derivatised sugars by PAGE or CZE.
Identifying Sites of Glycosylation in Proteins
As described above, the routine analysis of glycoproteins usually involves the analysis of the pooled oligosaccharides released from the protein and rarely are specific sites of glycosylation assigned. This leads to the loss of positional information such as the identification of the in vivo acceptor site specificity of a particular glycosyltransferase, whether there is site specific heterogeneity and how much sugar is at each site of glycosylation.
Mass spectrometry (MS) has played an important role in improving the approach to oligosaccharide analysis of glycoproteins and glycopeptides, particularly electrospray ionization MS (ESI-MS). Liquid chromatography coupled 'in-line' to an ESI-MS has proved to be a powerful tool for the rapid isolation and characterization of glycopeptides and glycoproteins. Under the right conditions, carbohydrate specific-ions for HexNAc-Hex, HexNAc, Hex and NeuAc can be generated in the mass spectrometer which allow the rapid identification of a glycoprotein or glycopeptide (Carr et al, 1993). However, since the method involves identification using a mass detector, it is not possible to determine which hexoses and hexosamines are in a particular carbohydrate (eg; GlcNAc and GalNAc cannot be distinguished). Mass spectrometry also lacks the capacity to specifically identify individual sites of glycosylation in clustered O-glycosylation domains, such as those found in mucins and the 'mucin-like' domains on many cell surface glycoproteins (Carraway and Hull, 1991).
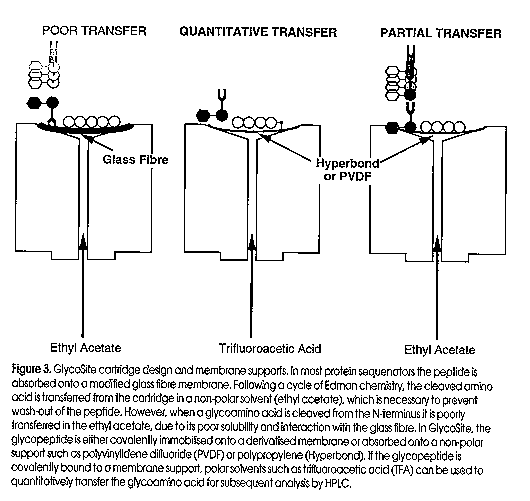
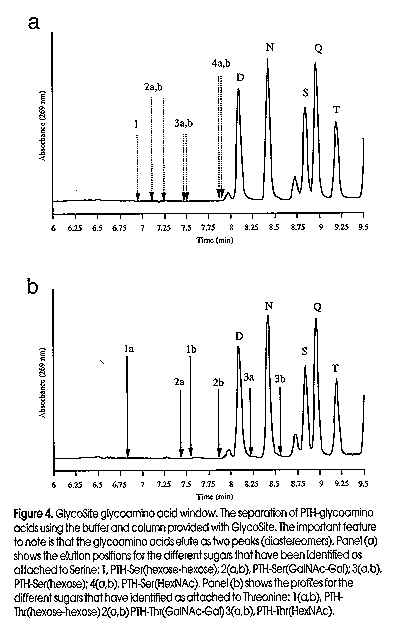
An approach we have pioneered for the identification of site-specific glycosylation is solid-phase Edman degradation, a chemical method where glycosylated amino acids are recovered as phenylthiohydantoin (PTH)-glycoamino acids during routine protein sequencing (Gooley et al, 1995). In July a protein sequencer dedicated to glycosylation site identification, GlycoSite , will be released by Beckman Instryments Inc. GlycoSite has been developed with the covalent attachment chemistries and chromatography suitable for the identification of all three major glycosylated amino acids, PTH-Asn(Sac), PTH-Thr(Sac) and PTH-Ser(Sac). Prior to GlycoSite , the typical protein sequencer used a modified glass fibre membrane to absorb the peptide/protein (Figure 3). Following cleavage from the protein, most of the glycoamino acid remains absorbed to the glass fibre and only a small quantity of glycoamino acid is transferred to an HPLC for analysis. In the GlycoSite , the glycopeptide is covalently immobilised onto a membrane support which allows the use of a more polar solvent, such as liquid trifluoroacetic acid (TFA), to transfer the glycoamino acid to the HPLC. Partial transfer of small chain length glycoamino acids is also possible if the glass fibre support is exchanged for non-polar membrane supports such as PVDF, Teflon and Hyperbond (Figure 3, Pisano et al, in press). GlycoSite has been used to characterize site specific glycosylation from a variety of eukaryotes, including mammals, fungi and the cellular slime mold Dictyostelium discoideum (Figure 4). In addition, sufficient amounts of PTH-glycoamino acid have been recovered by solid-phase Edman degradation for mass analysis by ESI-MS and compositional analysis by HPAEC PAD (Gooley et al, 1994). GlycoSite is the best technology for glycosylation site identification, allowing quantitative recovery of glycoforms and separates the three common glycoamino acids, Asn(Sac), Ser(Sac) and Thr(Sac) (Figure 4).
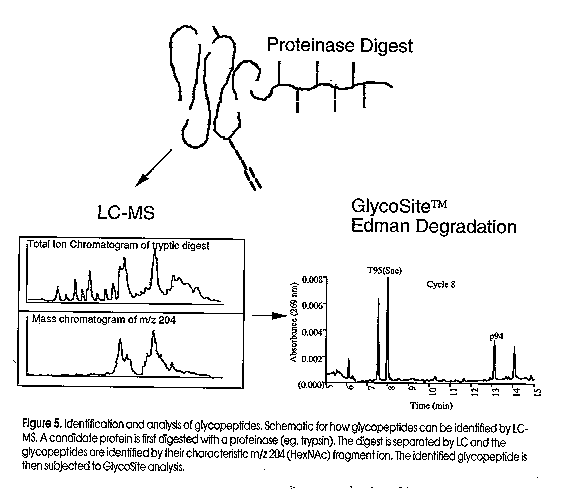
The combined approach of LC-ESI MS and solid-phase Edman degradation provides a powerful approach to rapidly identify and recover glycopeptides and glycoamino acids (Figure 5). The candidate glycoprotein is digested with a specific proteinase and the resulting peptides are subjected to LC-ESI MS analysis. Glycopeptides are identified by their characteristic carbohydrate specific-ions and collected for GlycoSite analysis. The positive identification of glycosylation sites allows the protein chemist to study the acceptor site specificity of any glycosyltransferase in vivo. This will prove a powerful new strategy for the identification of new glycosyltransferases, particularly in the simpler eukaryotes which are increasing in popularity as expression systems for recombinant proteins. With the development of MS compatible Edman reagents (Aebersold et al, 199X), the future looks very bright for high sensitivity analysis of any glycoprotein. Figure Legends:
Figure 2. State-of-the-Art technology used in the analysis of glycoproteins. The four boxes shown represent the key steps in oligosaccharide analysis, namely release from the protein, derivatisation, separation and detection. Two companies have released derivatisation kits (box 2) for the reductive amination of released oligosaccharides. Oxford Glycosystems (OGS) have developed amino benzamide (AB) for HPLC separation and aminobenzoic acid (AA) for electrophoresis. Glyko, and more recently Bio-Rad and Beckman Instruments, market amino naphthalene trisulfonic acid (ANTS) for the electrophoretic separarion of labelled oligosaccharides. MUCAB has made innovations in two areas of the above technologies; namely new chemistries for derivatisation of the oligosaccharides (International Publication No. WO 95/15969) and desalting (patent applied for). Figure 3. GlycoSite cartridge design and membrane supports. In most protein sequenators the peptide is absorbed onto a modified glass fibre membrane. Following a cycle of Edman chemistry, the cleaved amino acid is transferred from the cartridge in a non-polar solvent (ethyl acetate) which is necessary to prevent wash-out of the peptide. However when a glycoamino acid is cleaved from the N- terminus it is poorly transferred in the ethyl acetate, due to its poor solubility and interaction with the glass fibre. In GlycoSite, the glycopeptide is either covalently immobilised onto a derivatised membrane or absorbed onto a non-polar support such as polyvinylidene difluoride (PVDF) or polypropylene (Hyperbond). If the glycopeptide is covalently bound to the membrane support, polar solvents such as trifluoroacetic acid (TFA) can be used to quantitatively transfer the gllycoamino acid for subsequent analysis by HPLC. Figure 4. GlycoSite glycoamino acid window. The separation of PTH-glycoamino acids using the buffer and column provided with GlycoSite. The important feature to note is that the glycoamino acids elute as two peaks (diastereomers). Panel (a) shows the elution positions for the different sugars that have been identified as attached to serine; 1) PTH-Ser(hexose-hexose) 2a,b) PTH-Ser(GalNAc-Gal) 3a,b) PTH-Ser(hexose) 4a,b) PTH-Ser(HexNAc) Panel (B) shows the profiles for the different sugars we have identified attached to Threonine: 1a,b) PTH-Thr(hexose-hexose) 2a,b) PTH-Thr(GalNAc-Gal) 3a,b) PTH-Thr(HexNAc) Figure 5. Identification and analysis of glycopeptides. Schematic for how glycopeptides can be identified by LC-MS. A caandidate protein is first digested with a proteinase (eg. trypsin). The digest is separated by LC and the glycopeptides are identified by their characteristic m/z 204 (HexNAc) fragment ion. The identified glycopeptide is then subjected to GlycoSite analysis.
Glycoprotein research in MUCAB has been supported by the ARC and NH&MRC grants to AAG, NHP, JWR and KLW. GlycoSite was a joint project between MUCAB and Beckman Instruments Inc.
References
Aebersold, R., Bures, E.J., Namchuk, M., Goghari, M.H., Shushan, B. and Covey, T.C. (1992) Prot. Sci. 1, 494-503.
Carr,S.A., Huddleston,M.J. and Bean,M.F. (1993) Prot. Sci., 2, 183 196.
Carraway,K.L. and Hull,S.R. (1991) Glycobiology, 1, 131 138.
Dwek,R.A., Edge,C.J., Harvey,D.J., Wormald,M.R. and Parekh,R.B. (1993) Ann. Rev. Biochem. 62, 65-100.
Gooley,A.A., Pisano,A., Packer,N.H., Redmond,J.W., Williams,K.L (1994) Glycoconjugate J., 11, 180 186.
Gooley, A.A. and Williams, K.L. (1994) Glycobiology, 4, 413.
Gooley, A.A., Packer, N.H., Pisano, A., Redmond, J.W., Alewood, P.F., Jones, A., Loughnan, M., Williams, K.L. In: Crabb, J.W. ed. (1995) Techniques in Protein Chemistry VI., Academic Press, Inc. USA, pp. 83 90.
Hardy, M.R. and Townsend, R.R. (1994) In: Guide to techniques in glycobiology, Lennarz, W.J. and Hart, G.W. (Ed). Meth. Enzymol., 230, 208-225.
Packer, N.H., Wilkins, M.R., Golaz, O., Lawson, M., Gooley, A.A., Hochstrasser, D.F., Redmond, J.W. and Williams, K.L. (1996) Bio/Technology, 14, 66-70.
Patel, T.P. and Parekh, R.B. (1994) Release of oligosaccharides from glycoproteins by hydrazinolysis. In: Guide to techniques in glycobiology, Lennarz, W.J. and Hart, G.W. (ed). Meth. Enzymol., 230, 57-66.
Varki,A. (1993) Glycobiology 3, 97 130. Copyright 1996 Australian Biotechnology Association Ltd. The following images related to this document are available:Line drawing images[au96003c.gif] [au96003a.gif] [au96003e.gif] [au96003b.gif] [au96003d.gif] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}