 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Volume 8 Number 1, January/February 1998, pp. 47-49
Molecular Ecology of the Microbial Degradation of Aromatic CompoundsGareth Lloyd-Jones, Andrew D. Laurie, and David W.F. Hunter Landcare
Research, Private Bag 3127, Hamilton, New Zealand
Code Number:AU98012
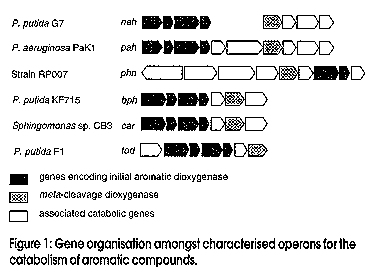
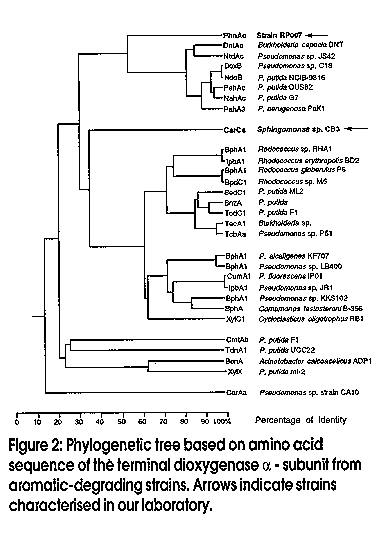
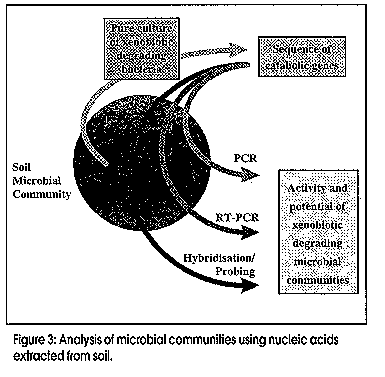
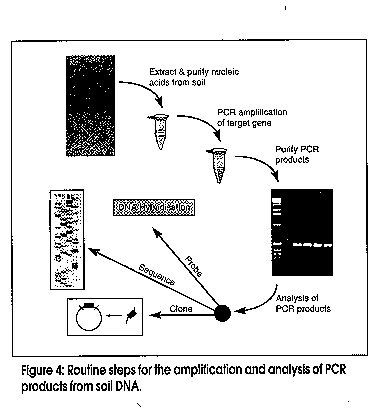
Microbes can be recognized by direct analysis of nucleic acids without the unrepresentative and unreliable steps of classical cultivation by using the techniques of molecular biology. This paradigm shift in how we conduct environmental microbiological research is relatively recent, most of the research and methods having originated during the past decade. The approach has great potential to complement chemical and physical methods for evaluating the potential for bioremediation, and subsequent monitoring of progress. Microbes have been recognised for more than a century as the primary agents in the ecology of carbon flux in nature. Research effort spanning decades has revealed that a wide variety of microorganisms are capable of degrading an equally wide range of organic pollutants (Aislabie and Lloyd-Jones, 1995; Lloyd-Jones and Hunter, 1997; Aislabie et al., 1997). Organic matter, as well as organic contaminants - e.g., petroleum products, solvents, lubricants, industrial process chemicals and pesticides - are gradually removed from soils by these natural microbial processes. Removal rates may be very slow for the more recalcitrant compounds, which include polycyclic aromatic hydrocarbons (PAHs), nitrogen heterocycles, polychlorinated biphenyls (PCBs), organochlorine pesticides, and pentachlorophenol. This natural removal of contaminants has been termed `intrinsic bioremediation' and is the process which is enhanced when bioremediation is applied as a clean-up option to contaminated sites. Bacterial enzyme systems provide the biological mechanism whereby organic contaminants are removed from polluted environments. An essential class of enzymes involved in the aerobic degradation of organic pollutants are the oxygenases. These catalyse reactions in which atoms of oxygen are incorporated into organic substrates, and are classified in two groups: (i) dioxygenases that catalyse the incorporation of both atoms of dioxygen into substrates; (ii) monooxygenases that catalyse the insertion of one atom from dioxygen. The enzymic reactions involved in the oxidation of aromatic hydrocarbons such as phenanthrene, naphthalene, biphenyl, and toluene, which are ubiquitous environmental pollutants, follow common paths. The first reaction in aromatic catabolic pathway involves the introduction of two hydroxyl groups into an aromatic ring by an aromatic ring dioxygenase. Subsequent reactions lead to cleaving of the aromatic ring by either an extradiol (meta-cleavage) or an intradiol (ortho-cleavage) ring-cleavage dioxygenase. Many genes which encode for these dioxygenases and associated enzymes required for the degradation of different aromatic compounds have been characterised in detail. Catabolic genes from different bacteria are often arranged in discrete clusters, or operons, and the corresponding nucleotide sequences are deposited in databases such as Genbank and EMBL. Our studies with bacteria isolated from contaminated sites in New Zealand have revealed a high degree of similarity in the specific degradative genes between these organisms and those from comparable bacterial isolates from the USA, Japan, and Antarctica (Lloyd-Jones and Lau, 1997). During these studies we have elucidated the sequences of two bacterial operons, one involved in the degradation of the azaarene carbazole (Shepherd and Lloyd-Jones, 1997), and the other involved in the degradation of the PAH phenanthrene. Comparison of the catabolic genes from these and other operons reveals many common patterns. Operons which encode for enzymes that catalyse the degradation of aromatic compounds often show similar patterns in their gene arrangement (Figure 1). The gene designations refer to the aromatic compound that is degraded by the genes of the operon: hence nah - naphthalene, pah/phn - phenanthrene, bph - biphenyl, car - carbazole, and tod - toluene. At the level of individual genes we also see conserved patterns within similar gene families whereby the predicted amino acid sequences reflect enzyme function. Many of these genes form distinct clusters of closely related sequences, which are revealed by phylogenetic analysis. The phylogenetic tree (Figure 2) shows a comparison of the terminal dioxygenase a-subunit derived from a number of aromatic-degrading strains. The terminal dioxygenase a-subunit constitutes part of the aromatic compound dioxygenase, and confers substrate specificity to the enzyme. The phylogenetic groupings reflect the aromatic substrate specificity of the enzymes. On the basis of these distinct similarities and differences between isofunctional genes we are able to design gene probes that are specific for the degradation of selected aromatic compounds. A closer inspection of the subgroups in Figure 2 reveals distinct classes, e.g., those specific for PAHs such as naphthalene and phenanthrene, or biphenyl/PCBs. Figure 1: Gene organisation amongst characterised operons for the catabolism of aromatic compounds Figure 2: Phylogenetic tree Analysis of the distribution of these essential enzymes of detoxification will prove useful in determining the response of microbes to the challenge offered by organic contaminants. The distribution provides a measure of genotypic diversity, which is the genetic potential (abundance and diversity) within a microbial population. However, only with prior knowledge of specific gene sequences are we able to apply this information to our search to understand what is occuring within the soil environment (Figure 3). Studies of genotypic diversity allow us to answer such questions as: How representative are characterised sequences of the natural microbial population? How broadly are these genes distributed? Are degradative genes concentrated where contaminant levels are diluted and not toxic, or present where contaminants are concentrated? Figure 3: Analysis of microbial communities using nucleic acidsmextracted from soil Before this can be achieved we must be able to reproducibly isolate nucleic acids directly from all soils, and further, this nucleic acid must be representative of the sampled population. Purification of nucleic acids from soils involves extraction of the DNA from intact microbial cells and procedures for the removal of organic contaminants such as humic acids, which can interfere with downstream analyses. We are in the process of validating a range of extraction/purification methods appropriate for a range of New Zealand soils. It is important to realise, however, that there will be no single method of nucleic acid purification that will be suitable for all soils. Contaminated soil and sediment samples can be studied to determine the distribution of genes encoding enzymes involved in the degradation of specific contaminants. This can be achieved using a PCR-based probing strategy targeting DNA extracted from soils (Figure 4). The high specificity, sensitivity, and reproducible consistency of PCR detection of specific DNA sequences from complex environmental samples, though still in its infancy, will contribute enormously to our understanding of environmental microbiology. The PCR fragments shown in Figure 4 illustrate the amplification of degradative genes from soils severely contaminated with PAHs (Lloyd-Jones and Lau, 1997). Figure 4: Routine steps for the amplification and analysis of PCR products from soil DNA Leading on from such studies, we need to further clarify the ecological significance of specific genes. This phenotypic diversity refers to the expression of genes under given conditions which cannot be determined until the extent of the in situ expression of these genes is demonstrated. Several recent studies have suggested that hybridisation to target mRNA may be used as a means to monitor the expression of specific genes. The increased sensitivity of transcript detection using reverse transcriptase PCR may provide for improved detection of these mRNA transcripts. Since Taq DNA polymerase will not amplify from transcribed mRNA, the viral enzyme reverse transcriptase is used to effect cDNA synthesis from isolated mRNA. This cDNA is subjected to PCR amplification with primers specific for the transcribed mRNA that is the target of the experiment. Conclusion The presence of specific microbial genes and their expression in response to contamination are potentially useful for: (i) determining whether our current sequence databases adequately reflect what occurs in natural microbial communities; (ii) predicting whether pollutants are present at levels that affect soil sustainability; (iii) contributing to risk assessment models for ranking soils to identify those requiring immediate remediation; and (iv) evaluating the potential for bioremediation and for monitoring its in situ progress. References Aislabie, J., Hunter, D.W.F., Lloyd-Jones, G. 1997. Polyaromatic compound-degrading bacteria from a contaminated site in New Zealand. In situ and On-site Bioremediation 4(2):219-224. Aislabie, J., Lloyd-Jones, G. 1995. Bacterial degradation of pesticides. Australian Journal of Soil Research 33:925-942. Lloyd-Jones, G., Hunter, D.W.F. 1997. Characterization of fluoranthene and pyrene-degrading mycobacterium-like strains by RAPD and SSU sequencing. FEMS Microbiology Letters 153:51-56. Lloyd-Jones, G., Lau, P.C.K. 1997. Glutathione s-transferase-encoding gene as a potential probe for environmental isolates capable of degrading polycyclic aromatic hydrocarbons. Applied and Environmental Microbiology 63:32863290. Shepherd, J.M., Lloyd-Jones, G. Identification of a gene cluster expressed during growth of Sphingomonas CB3 on carbazole. Gene (submitted). Copyright 1998 Australian Biotechnology Association Ltd. The following images related to this document are available:Line drawing images[au98012d.gif] [au98012b.gif] [au98012a.gif] [au98012c.gif] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}