 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Volume 8 Number 2, March/April 1998, pp. 86-90 Special Feature Vaccine Biotechnology
Code Number:AU98016
Our Special Feature for this issue highlights developments in Vaccine Biotechnology. We are fortunate indeed to have several contributions from members of the Cooperative Research Centre for Vaccine Technology led by Professor Michael Good of the Queensland Institute of Medical Research (QIMR). The first paper in our collection is from Dr Richard Strugnell's group (CRC Vaccine Research/University of Melbourne), and it deals with the vexing question of why bacterial vectors have failed to deliver the goods. Next, Dr Robin Anders and co-workers (CRC Vaccine Research/The Walter and Eliza Hall Institute) describe a leading malaria vaccine candidate. Dr David Boyle (CSIRO/AAHL) then describes a relatively new concept for deploying new combinations of priming and boost antigenic exposures which is showing potential for improving immune responses to novel "recombinant" vaccines. We have two additional papers from CSL Ltd on DNA Vaccines and Adjuvant technology which we plan to include in our next journal issue to complete this series on Vaccinology. Thanks to all the participants for these contributions, and we trust the biotech community will find this feature a useful update on the vaccine field. David Tribe, Bacterial Vectors: why have they failed to deliver ? Richard Strugnell, Cameron Simmons, Odilia Wijburg, Tania Uren, Damien Drew and Sarah Dunstan, Department of Microbiology and Immunology, University of Melbourne, Parkville, VIC 3052 r.strugnell@microbiology.unimelb. edu.au Bacterial vaccine vectors have been studied experimentally for over twenty years
yet there are
no vector-based vaccines in commercial human or veterinary use today. The absence of
human vector-based vaccines is probably due to problems associated with testing human
vaccines, and a lack of funds to support vaccines for use in developing countries.
Vector-based veterinary vaccines are currently not competitive with combination adjuvanted
vaccines but may have a niche under certain circumstances. The future of vaccine vectors
may lie in the delivery of DNA vaccines.
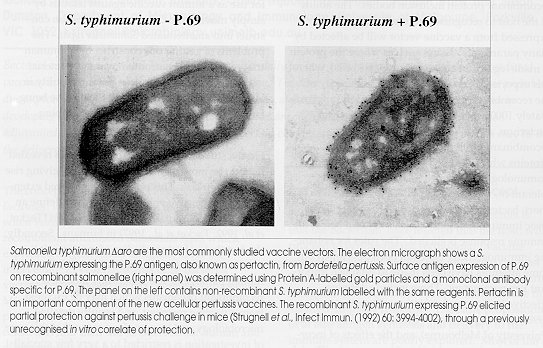
The answer to this question in the human context is complex and must accommodate issues including the availability of human vaccine testing facilities, support for new vaccines relevant to developing countries, and vaccine regulatory issues. The reasons why bacterial vectors have not assumed a strong position in veterinary preventative medicine are less clear but efficacy, the intensity of the vectored response, the conservative nature of veterinary vaccine manufacture which has evolved from the low profit margins of veterinary vaccines, and the negative perception associated with injecting live, albeit attenuated, pathogens into otherwise healthy farm animals, are all central. Some of these issues we have identified which relate to bacterial vaccine research and development will be examined further in this article. This article will not address vaccine delivery from recombinant normal bacterial flora since the capacity of such vectors to deliver a range of antigens to different species has not been demonstrated. The Bacterial Vaccine Vector Concept The basic premise of bacterial vaccine vectors is that the bacterium, once injected into the host, will proceed to synthesise the recombinant vaccine antigen in vivo and deliver this protein to the host immune system. The immune system will, in turn recognise the recombinant protein and produce the `desired' immune response. The basic propositions of in vivo antigen synthesis and delivery to the immune system are poorly explored. Most of the bacterial vaccine vector studies have been performed in mutant Salmonella typhimurium which are attenuated by mutation of the aromatic synthesis pathway (ie. Daro)(Hoiseth & Stocker, 1981). This bacterium was originally constructed as a vaccine rather than a vaccine vector. Is it sensible to suggest that a bacterium which is unable to carry out de novo synthesis of aromatic compounds, including aromatic amino acids, will be an ideal `factory' for producing recombinant antigens in vivo ? Dunstan et al. (to be submitted) used sensitive reporter systems such as firefly luciferase to examine in vivo gene expression from the paradigm vector Salmonella typhimurium Daro, in vivo in isolated Peyer's patches from immunised animals. They determined that regulated and constitutive bacterial promoters which have different capacities to express luciferase in Peyer's patches, have different capacities to elicit immune responses when used to drive expression of candidate vaccine antigens. A correlation between absolute expression level and immune response was not observed, suggesting that an optimal level of expression may be one which is less than maximal. This makes sense if it is important to preserve a level of virulence in bacterial vector, a level which may be reduced if the bacterium is largely full of recombinant protein inclusion bodies. The ability of the host to recognise a recombinant protein expressed from a vaccine vector will be affected by many parameters. Issues such as where the protein is made (eg. Salmonellae reside inside cells), where it is exported to, the intrinsic immunogenicity of the recombinant protein versus that of the approximately 1000 other gene products of the carrier bacterium, and the similarity between the recombinant protein and other normal flora proteins which the host may have been immunologically primed against or rendered tolerant to, will all impact on immunogenicity. In short, bacterial vaccine vectors have a number of basic immunobiological hurdles to jump before an immune response is elicited. The rules for enhancing immunogenicity have not been developed but some attempts have been made to establish important key parameters. These include defining effective background mutations (Dunstan et al., 1998), gene copy number (Dunstan SJ, Ph.D thesis, University of Melbourne), and the effects of incorporating cytokine genes into the constructs (Denich et al., 1993; Dunstan et al., 1996) on the immunogenicity of recombinant vectors. The Particular Problems of Vaccine Vectors in Humans There are numerous examples of experimental bacterial vaccine vectors which have been successful in murine systems despite the problems identified above. Perhaps the most effective example of a bacterial vectored vaccine is the oral tetanus vaccine developed by Fairweather et al. (1990) which comprised a S. typhimurium Daro expressing the non-toxic binding portion of tetanus toxin known as the C fragment. This vaccine elicits high titre (ie. >100,000), long lasting and complete antibody-mediated protection against tetanus toxin challenge in mice. Studies by Fairweather et al. showed that the vaccine was safe in the rodent model and, theoretically, could be readily adapted for use as a human vaccine against tetanus by simply moving the C fragment expression construct into an attenuated Salmonella typhi. The problems of getting this construct into a human trial reflect those confronted by many bacterial vaccine vectors which a have potential utility in humans, and especially where the disease being targeted is predominantly one of developing countries. Firstly, studies of S. typhi Daro in humans revealed them to be more virulent than expected, giving rise to `vaccinemias'. This observation required extensive and expensive volunteer studies to define an optimal carrier mutation for use in S. typhi (Tacket et al, 1992; Tacket et al., 1997) in humans. Secondly, the fact that volunteer studies of most vaccine vectors examine the virulence of potentially lethal human pathogens, use genetically modified recombinant organisms which may be excreted from a vaccine recipient, and require detailed microbiology and immunology mean that this type of investigation is restricted to a very few specialist facilities such as the Center for Vaccine Development (CVD) in Baltimore, USA. Thirdly, the development of a non-parenteral (ie S. typhi-based) tetanus vaccine, though identified by the WHO as an important milestone in vaccine development (http://www.who.ch/gpv/), may have a relatively small commercial market and might predominantly be used in developing countries where neonatal tetanus and typhoid remain a problem. There is therefore limited commercial support for the development of such a vaccine. It is testament to the energy of the CVD director, Myron Levine, that substantial progress has been made towards the testing of the oral tetanus/typhoid vaccine in humans. The position of the regulatory authorities, who increasingly demand more exact definitions of vaccine components, on live attenuated recombinant bacterial pathogen-based vaccines has not been thoroughly tested and quality control of such vaccines may be both technically complex and expensive. While it may be relatively simple for say a bacterial vector-based HIV vaccine to obtain registration, the prospects of replacing an existing highly efficacious vaccine (such as tetanus toxoid) under some circumstances with a new live vector-based vaccine are less rosy. The fact that live attenuated typhoid vaccines will be separately tested and developed for use in Western travellers and typhoid endemic countries, and that Mycobacterium bovis strain BCG has been thoroughly tested in humans as a vaccine (Cohn, 1997), may make these attenuated S. typhi and BCG more attractive to regulatory authorities as vehicles for delivering recombinant antigens. Veterinary Vaccine Vectors Figure: Salmonella typhimurium Some of these problems identified with bacterial vaccine vectors in humans are less important in veterinary medicine. In the development of veterinary vaccines it is usual for the vaccines to be investigated in the target species and problems associated with extrapolation of results across species do not then arise. Veterinary vaccination is primarily driven by considerations of cost - the cost of the disease versus the cost of the vaccine. While live attenuated bacterial vaccines which can be produced by fermentation, then lyophilised, appear to be potentially highly cost-effective, the addtional costs of quality control might erode the small profit margins available to the veterinary vaccine manufacturer. Such costs could include those involved in determining stability of antigen expression by the vector, maintaining shelf life, and identifying and controlling a vaccine dose which avoids site reactions and systemic sequelae such as disseminated infection by the vector, coupled with a requirement, where possible, for single dose efficacy, and may mean that a vector-based veterinary vaccine would be subject to very tight product specifications. It is sometimes said that the rural sector is conservative and that this propensity can inhibit the development of new technologies. While this may be true on occasion, the perception problem associated with the infection of healthy animals with attenuated pathogens expressing vaccine antigens from other organisms is real and must be addressed. Animal producers and, more importantly, consumers will have to be thoroughly convinced that it is both safe and necessary to infect a healthy pig, sheep or cow with a "pathogenic" organism such as a Salmonella species in order to stop it becoming infected with another microorganism which may not carry the same stigma. The major reason, however, for the absence of veterinary vaccine vectors in commercial use must be efficacy and intensity of response. High efficacy (ie. >80%), which may be simple to demonstrate in experimentally vaccinated inbred mice housed under laboratory conditions, may fall away when the vaccine is used in outbred animals which are subject to nutritional or other environmental stresses. Potent adjuvants, deemed too toxic for human use, are effective at eliciting protective responses from killed organisms or from subunit preparations in animals. Thus, apart from poultry where margins demand inhalation or ingestion of vaccines, it is difficult to see the responses elicited by live attenuated bacteria being competitive with those elicited by killed adjuvanted vaccines. The diseases for which vaccines do not exist but for which there are strong commercial markets (eg. the gastrointestinal nematodes of sheep), include many infections for which correlates of protection are yet to be defined. Identification of correlates which are consistent with the types of responses elicited by live recombinant bacterial vaccines (eg. T-helper type 1 (TH1) responses induced by Mycobacterium species (Lagranderie et al., 1996)) may lead to the development of vaccine vectors for singular and specific applications. It is unlikely however that vaccine vector technology will become generic and replace the relatively inexpensive, single dose adjuvanted combination vaccines favoured in extensive animal husbandry today. Whither Vaccine Vectors The lack of commercial success with vaccine vectors has almost certainly slowed research
and
development of the technology. This commercial reality does not sit well with those who
know
that `life-long' protection is only achieved following infection with pathogens or following
administration of live vaccines. This observation coupled with the efficacy problems of existing
typhoid and tuberculosis vaccines suggest that live, genetically and rationally attenuated
S.typhi and M. tuberculosis will remain an option for new
improved human vaccines against
these important diseases. If and when such vaccines are fully developed, the opportunity will
arise for them to act as vectors for other heterologous `protective' antigens. In the veterinary
field, a niche may open for the technology if a specific application, that is The biggest threat, and perhaps the biggest opportunity, for vaccine vector research may be in the area of DNA vaccination. Unmodified DNA vaccination elicits similar types of responses to those seen with the intracellular bacterial vaccine vectors eg. Salmonellae and Mycobacteria with cytotoxic T cells, TH1 and humoral immunity (Ulmer et al., 1997). However, significant problems exist with the delivery of DNA vaccines. Intramuscular injection of large amounts of DNA results in relatively inefficient in vivo transfection, particularly in larger animals, and the high level responses obtained in murine systems from single injections of DNA have not generally been observed in vaccination trials on larger animals. A recent report by Darji et al. (1997) suggests that vaccine vectors are not limited to the delivery of protein antigens but also may efficiently deliver DNA vaccines. This observation, and previous studies by Sizemore et al. (1995, 1997) who showed that recombinant Shigellae could deliver DNA vaccine-encoded reporter genes to the mucosal epithelium suggest that vaccine vectors hold some promise in this regard. The use of a bacterial vaccine vector as both the means of producing and delivering a DNA vaccine has intrinsic logic and studies are under way in a number of centres (Pascual et al., 1997) to examine the capacity of bacterial vaccine vectors to deliver DNA vaccines. RS, CS and TU are members of the CRC for Vaccine Technology. References Chatfield SN, Strugnell RA, Dougan G. (1989) Live Salmonella as vaccines and carriers of foreign antigenic determinants. Vaccine 7:495-498. Cohn DL. (1997) Use of the bacille Calmette-Guerin vaccination for the prevention of tuberculosis: renewed interest in an old vaccine. Am J Med Sci 313:372-376. Darji A, Guzman CA, Gerstel B, Wachholz P, Timmis KN, Wehland J, Chakraborty T, Weiss S. (1997) Oral somatic transgene vaccination using attenuated S. typhimurium. Cell 91:765-775. Denich K, Borlin P, O'Hanley PD, Howard M, Heath AW. (1993) Expression of the murine interleukin-4 gene in an attenuated aroA strain of Salmonella typhimurium: persistence and immune response in BALB/c mice and vaccine susceptibility to macrophage killing. Infect Immun 61:4818-4827. Dunstan SJ, Ramsay AJ, Strugnell RA. (1996) Studies of immunity and bacterial invasiveness in mice given a recombinant salmonella vector encoding murine interleukin-6. Infect Immun 64:2730-2736. Dunstan SJ, Simmons CP, Strugnell RA. (1998) Comparison of the abilities of different attenuated Salmonella typhimurium strains to elicit humoral immune responses against a heterologous antigen. Infect Immun 66:732-740. Dunstan SJ, Simmons CP, Strugnell RA. Use of in vivo regulated promoters in the delivery of antigens from Salmonella enterica var. Typhimurium. (to be submitted). Fairweather NF, Chatfield SN, Makoff AJ, Strugnell RA, Bester J, Maskell DJ, Dougan G. (1990) Oral vaccination of mice against tetanus by use of a live attenuated Salmonella carrier. Infect Immun 58:1323-1326. Formal SB, Baron LS, Kopecko DJ, Washington O, Powell C, Life CA. (1981) Construction of a potential bivalent vaccine strain: introduction of Shigella sonnei form I antigen genes into the galE Salmonella typhi Ty21a typhoid vaccine strain. Infect Immun 34:746-750. Hodgson ALM. (1994) Bacterial vaccine vectors, p.177-186. In PR Wood, P Willadsen, JE Vercoe, RM Hoskinson, and D. Demeyer (eds). "Vaccines in Agriculture: Immunologicals Applications to Animal Health and Production". CSIRO, Australia, Melbourne. Hoiseth SK, Stocker BAD. (1981) Aromatic-dependent Salmonella typhimurium are non-virulent and effective as live vaccines. Nature 291:238-239. Lagranderie MR, Balazuc AM, Deriaud E, Leclerc CD, Gheorghiu M. (1996) Comparison of immune responses of mice immunized with five different Mycobacterium bovis BCG vaccine strains. Infect Immun 64:1-9. Pascual DW, Powell RJ, Lewis GK, Hone DM. (1997) Oral bacterial vaccine vectors for the delivery of subunit and nucleic acid vaccines to the organized lymphoid tissue of the intestine. Behring Inst Mitt 98:143-152. Sizemore DR, Branstrom AA, Sadoff JC. (1995) Attenuated bacteria as a DNA delivery vehicle for DNA-mediated immunization. Science 270:299-302. Sizemore DR, Branstrom AA, Sadoff JC. (1997) Attenuated Shigella as a DNA delivery vehicle for DNA-mediated immunization. Vaccine 15:804-807. Tacket CO, Hone DM, Curtiss R 3d, Kelly SM, Losonsky G, Guers L, Harris AM, Edelman R, Levine MM. (1992) Comparison of the safety and immunogenicity of DaroC DaroD and DcyaDcrp Salmonella typhi strains in adult volunteers. Infect Immun 60:536-541 Tacket CO, Sztein MB, Losonsky GA, Wasserman SS, Nataro JP, Edelman R, Pickard D, Dougan G, Chatfield SN, Levine MM. (1997) Safety of live oral Salmonella typhi vaccine strains with deletions in htrA and aroC aroD and immune response in humans. Infect Immun 65:452-456. Ulmer JB, Deck RR, DeWitt CM, Donnelly JJ, Friedman A, Montgomery DL, Yawman AM, Orme IM, Denis O, Content J, Huygen K, Liu MA. (1997) Induction of immunity by DNA vaccination: application to influenza and tuberculosis. Behring Inst Mitt 98:79-86. Copyright 1998 Australian Biotechnology Association Ltd. The following images related to this document are available:Photo images[au98016a.jpg] |
| |||||||||
{kind=link}