 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
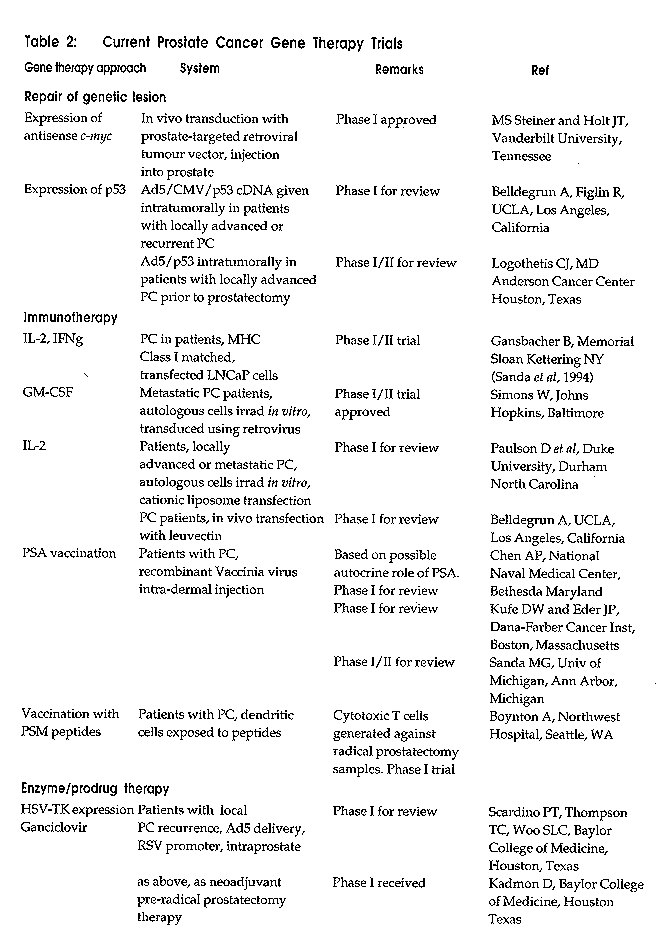
Volume 8 Number 2, March/April 1998, pp. 99-106 CONFERENCE PAPER Prostate Cancer Gene Therapy Russell PJ1, Martiniello-Wilks R1, Lockett LJ2, Brookes DE2, Zandvliet D2, Watt F2, Molloy PL2, Khatri A2, and Both GW2. 1Oncology Research Centre, Prince of Wales Hospital, NSW 2031 and
Division of Medicine,
University of New South Wales, NSW 2052, Australia;
Code Number:AU98019
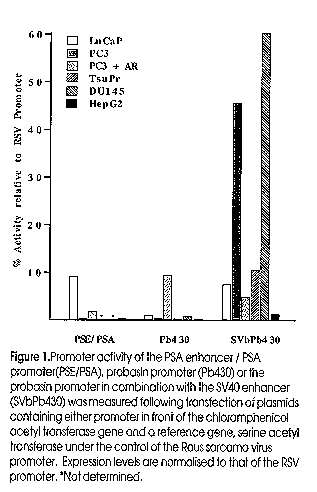
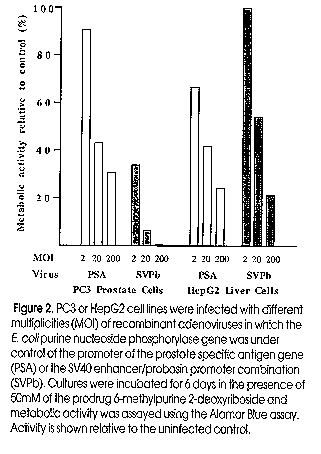
Prostate cancer is the most common cancer and the second most lethal cancer in men in the Western world. In Australia, each year around 15,000 new cases of prostate cancer appear and 2,600 Australian men die from this disease. In 1995, over 160,000 new cases were diagnosed in the US, and 45,000 men died, with associated health costs in excess of one billion dollars. Organ confined prostate cancer (PC) can be cured by surgery or by targeted irradiation, but control of cancer that has spread depends on the androgen requirement of prostate epithelial cells for both growth and survival. Treatments which suppress androgen synthesis and/or block its activity can lead to significant periods of remission, but androgen independent tumours, for which there is no effective cure, eventually emerge. Developing gene therapy technologies may provide new modalities for treating localised disease and patients with advanced prostate cancer. Strategies under consideration include both ex vivo and in vivo genetic modifications. Ex vivo strategies are based on manipulation of tumour cells in vitro to stimulate a host immune response, such as the use of GM-CSF-transduced prostate cancer vaccines, whilst in vivo strategies aim to overcome a genetic defect involved in PC progression (for example by inserting a tumour suppressor gene or by inducing apoptosis), or to express suicide genes in the target tumour cells. Gene Therapy Approaches for Cancer Genes can be delivered ex vivo or in vivo using physical methods or viral vectors (Sanda et al, 1994), and the choice of agent depends on the condition for treatment, efficiency of gene transfer, stability of gene expression and safety of clinical use. Unlike monogenic disorders, which require sustained expression or repeated delivery of a given gene, cancer should be amenable to treatment by transient expression and for this reason, adenovirus vectors are highly suitable. Human adenovirus 5 (Ad5) grow to high titers in HEK293 cells, infect a variety of dividing and nondividing cell types and provide good levels of transient gene expression (Vile and Russell 1994; Eastham et al, 1996). Several gene therapy strategies (Table 1) including the use of either replication-deficient Ad vectors to effect cell killing by EPT (Chen et al, 1995; Eastham et al, 1996) or to introduce a wild-type p53 gene (Ko et al, 1996), or replication competent Ads that are controlled by a tissue-specific promoter (Rodriguez et al, 1997) are under development. EPT relies on expression of a gene encoding an enzyme which activates a prodrug to a toxic metabolite that can spread to nearby cells creating a "bystander" effect. Not all cells need to express the enzyme to achieve comprehensive cell killing. Since the prostate is a dispensible organ, treatments which target cell killing to prostate epithelial cells, whether local or in metastases, should effect a cure. Conversion by HSVTK of the pro-drug, ganciclovir (GCV), to a toxic, phosphorylated form, a substrate for DNA synthesis, has been the most widely used system. Other EPT systems not dependent on DNA replication may be advantageous for PC and other tumours where the cell turnover rate is low (Berges et al, 1995). The E. coli enzyme, purine nucleoside phosphorylase (PNP), converts prodrugs such as 6-methylpurine deoxyriboside (6MPDR) or fludarabine (approved for clinical use) into toxic precursors, 6 methylpurine (6MP) or 2-fluoro-adenine, respectively, for both RNA as well as DNA synthesis (Sorscher et al, 1994). These compounds provide a highly efficient bystander effect because they are non-phosphorylated and can diffuse freely between cells. Even using a relatively weak promoter (PSA), recombinant adenovirus delivery of HSVTK or PNP was sufficient to inhibit the growth of PC3 human prostate cancer xenografts in nude mice and to prolong mouse survival (Martiniello-Wilks et al, submitted). Most of the above work relies on the ability to locally deliver the genes of interest to cancer cells, and many clinical trials of gene therapy for prostate cancer based on these studies are under review or have been approved (Table 2). Results from only one clinical trial are thus far available (trial by Scardino et al, Table 2 - , presented by Dr B Miles at Genitourinary Oncology Group Meeting, Leura, NSW, 1997). The safety of local injection of an EPT adenovirus into prostate was shown, but only limited clinical effects were seen. Apart from strategies which rely on an intervening immune response, these procedures are constrained to loco-regional injection into prostate tumours of patients under the guidance of transrectal ultrasound. Table 1: Development of gene therapy approaches using model systems. Table 2: Current prostate cancer gene therapy trials Targeting of tumour metastases Unlike melanomas and renal cell carcinomas which are immunogenic and occasionally display spontaneous tumour regression, PC is slow growing and poorly immunogenic, and spontaneous regression has not been observed (Hrouda and Dalgleish, 1996). However, the observation of tumour infiltrating lymphocytes (TIL) in PC suggests a possible role for these cells in tumour killing (Eastham et al, 1996; Ko et al, 1996). Moreover, TIL have been reported to be a good prognostic marker both for non-progression and survival in PC (Vesalainen et al, 1994). Candidate cells that mediate anti-tumour killing include cytotoxic T cells (CTL), activated macrophages and natural killer (NK) cells. Strategies which increase these cell numbers or activate their killing mechanisms have been used for immune gene therapy. For example, interleukin-2 (IL-2) is a potent growth factor and activator of CTL and NK cell tumour killing activity (Smith 1988; O'Shea and Ortaldo, 1992). Granulocyte-macrophage colony stimulating factor (GM-CSF) controls the growth differentiation and activation of macrophages (Gough et al, 1984). Interferon gamma (IFNg) enhances the expression of MHC class I and II molecules on a cell surface resulting in improved recognition and subsequently killing by CTLs. B7-1 is a molecule found on the surface of antigen presenting cells that binds CD28 on the surface of CTLs and NK cells providing an important co-stimulatory signal for the killing activity of these cells (Mueller et al, 1989; Nandi et al, 1994; Wu et al, 1995). Subsequently, studies have shown that the immune response to immunogenic as well as non-immunogenic tumour types may be amplified by the expression of B7-1 on the tumour cell surface in the presence of IL-2 (Garlepp et al, 1997). Studies in animal models have shown that genetic engineering of tumor cells to express particular cytokines can greatly enhance the immune response against the tumour, resulting in the reduction or complete inhibition of tumour progression and, in some cases, curing of established tumors. For example, the growth of Dunning MatLyLu prostate tumours was suppressed in rats inoculated with tumor cells transfected with IL-2 or GM-CSF (Vieweg et al, 1994). Other examples are described in Table 1. Experience with other tumor models e.g., colon cancer (Chen et al, 1995) suggests that cell killing combined with immune enhancement may be particularly effective. Thus, delivery and expression of cell killing and immune enhancing genes to an accessible tumour site should be able to effect elimination of both local and metastatic deposits. Alternative methods for targeting disseminated cancer cells (Hrouda and Dalgleish, 1996) include targeting of the neo-vascularisation required for nutrient supply and tumour growth, targeting of cytotoxic agents to tumour cells using antibodies or other ligands specific for the tumour cells, and targeting of recombinant therapeutic viruses using either such cell surface ligands or transcriptional specificity. Transcriptional targeting Transcriptional targeting involves the use of genetic elements to obtain gene expression under particular conditions or in specific cells. A recent example is the use of the human tyrosinase promoter/enhancer to control lacZ gene expression in melanoma cells (Siders et al, 1996). For prostate targeting, Henderson's group has used the enhancer and promoter of the prostate specfic antigen (PSA) gene to direct the expression of the E1A `master-switch' region in a replication-competent adenovirus, thus producing a replication competent virus with selectivity for cells expressing PSA (Rodriguez et al, 1997). Other candidate genes for useful regulatory elements include the rat probasin gene (androgen dependent) and the prostate specific membrane antigen (PSM) gene (androgen-independent). Some of our work in transcriptional targeting for prostate cells is presented below. Transcriptionally targeted vectors are most suitable when the virus can access the target cells directly e.g. by injection as this minimises losses due to infection of inappropriate cells. Strategies for systemic delivery of viruses to treat disseminated targets require cell surface targeting. Targeting of virus particles for systemic delivery Surface targeting of tumour cells by viruses potentially allows the systemic delivery of therapeutic agents to disseminated cancers. For efficacy, sufficient therapeutic agent must reach the target site without loss through binding to or infection of inappropriate tissues. The targeting agent must also exit the vascular system to penetrate the tumour. After binding to a surface marker, infection would occur via endocytosis mediated by Ad penton base-integrin interactions (Wickham et al, 1993). Systemic delivery to a surface marker requires that the normal tropism of Ad infection is blocked while infection of the target cells is enhanced in order to avoid extensive loss of virus due to inappropriate binding. `Proof of principle' was recently demonstrated in vitro (Douglas et al, 1996). The cell binding domain on the fiber protein of an Ad5 vector was modified using an antibody adaptor fragment chemically coupled to folate. Cells that expressed the folate receptor were preferentially infected, indicating that the adaptor blocked the normal route of uptake of the virus while creating a virus with a new cell binding specificity (Douglas et al, 1996). Prostate cells express a number of surface proteins which show a limited tissue distribution which could provide suitable targets, such as epidermal growth factor receptor (EGFR), luteinizing-hormone-releasing-hormone (LHRH) receptor, or insulin-like growth factor (IGF) receptors. A partcularly suitable antigen is prostate specific membrane antigen (PSM) (for review see (Israeli et al, 1997), which continues to be expressed in late stage PC (Israeli et al, 1994; Wright et al, 1996) and is elevated during androgen deprivation therapy. Whilst PSM is also expressed in the duodenum and small intestine, these should not be targeted by systemic delivery. Importantly, using anti-PSMA antibodies, intense staining is observed in endothelial cells or capillary vessels in peri- and endo-tumoral areas of malignancies including PC, renal cell carcinomas, transitional cell carcinomas and colon carcinomas, but not in normal vasculature, suggesting that PSM may be involved in tumour induced angiogenesis (Silver et al, 1997). Hence targeting to PSM should direct cell killing both the tumour vasculature as well as the tumour directly. For systemic delivery, pre-existing immunity to Ad5 may become an important consideration as natural human Ad infections are common. We have recently characterised and engineered a unique ovine adenovirus (OAV) as a vector (Vrati et al, 1996; Khatri et al, 1997; Xu et al, 1997); this virus is serotypically distinct and should not be neutralised by antibodies to human Ads offering an advantage in gene delivery. We have shown that OAV infects various human cell types, including prostate, PC3, and breast cancer cell lines, and can express a foreign gene. Importantly, OAV replication is abortive in human cells due to the absence of major late promoter function, as shown by RT-PCR analysis. In PC3 cells, at least one early viral promoter that has been examined also fails to function. It should be possible to use OAV vectors alone, or in tandem with human Ad vectors to deliver cell killing genes to tumors. Materials and Methods Transfections of reporter gene constructs and assays of expression were done using standard methods (Brookes et al, 1998). Expression cassettes were re-combined into replication defective human Ad5 (Lockett et al, 1997) and virus activity in cell killing determined by assaying cellular metabolic activity (Lockett et al, 1997). Results and Discussion Our group has worked extensively on producing targeted vectors containing prostate-specific promoters. The activity and specificity of the PSA, probasin (Pb) and relaxin H2 promoters was examined (Brookes et al, 1998). The Pb and PSA elements showed strong, androgen-inducible expression in prostate cells and limited expression in non-prostate cell types; both required active androgen receptor (AR) for activity. Studies by (Pang et al, 1997) have suggested that the complete PSA promoter contains two functional domains, a proximal promoter and a distal promoter which can also function as an enhancer, but which is also androgen responsive. These elements therefore may be of limited use for treating androgen-independent (AI) disease and could not be used in conjunction with common androgen ablation therapies. However, the combination of the SV40 enhancer with the Pb promoter (Sv/Pb) provided for high level expression in 3 AI PC cell lines and expression was not further increased by co-transfection of the AR ((Figure 1). Substantial specificity for prostate cells was observed, though significant expression was also seen in bladder and breast cell lines. When used to direct expression of an EPT gene in a recombinant Ad, cell killing was 5-10 fold greater in PC-3 prostate cells than in liver (HepG2) and lung fibroblast (MRC-5) cell lines in vitro (Figure 2). An equivalent ovine adenovirus carrying this expression cassette has been constructed. These viruses will allow us to compare the abilities of Ad5 and OAV vectors to deliver EPT to xenografted human tumours in a mouse model. Viruses of this type represent first generation tissue-specific vectors; there is substantial room for improvement and attempts to alter virus tropism at the cell surface have begun. Figure 1. Promoter activity of the PSA enhancer
Figure 2. PC3 or HepG2 cell lines infected with different multiplicities of recombinant adenoviruses References Asgari, K., Sesterhenn, I. A., McLeod, D. G., Cowan, K., Moul, J. W., Seth, P. and Srivastava, S. (1997) Inhibition of the growth of pre-established subcutaneous tumor nodules of human prostate cancer cells by single injection of the recombinant adenovirus p53 expression vector Int. J. Cancer, 71, 377-382. Berges, R. R., Vukanovic, J., Epstein, J. I., CarMichel, M., Cisek, L., Johnson, D. E., Veltri, R. W., Walsh, P. C. and Isaacs, J. T. (1995) Implication of cell kinetic changes during progression of human prostatic cancer Clin. Cancer Res., 1, 473-480. Brookes, D., Zandvliet, D., Watt, F., Russell, P. and Molloy, P. (1998) Relative activity and specificity of promoters from prostate-expressed genes The Prostate, In Press, Carducci, M. A., Ayyagari, S. R., Sanda, M. G. and Simons, J. W. (1995) Gene therapy for human prostate cancer: Translational research in the hormone refractory Dunning prostate cancer model Cancer, 75, 2013-2020. Chen, S.-H. C., Chen, X. H. L., Wang, Y., Kosai, K.-I., Finegold, M. J., Rich, S. J. and Woo, S. L. C. (1995) Combination gene therapy for liver metastasis of colon carcinoma in vivo Proc. Natl. Acad. Sci. USA, 92, 2577-2581. Dorai, T., Olsson, C., Katz, A. and Buttyan, R. (1997) Development of a hammerhead ribozyme against bcl-2. I. Preliminary evaluation of a potential gene therapeutic agent for hormone-refractory human prostate cancer Prostate, 32, 246-58. Douglas, J. T., Rogers, B. E., Rosenfeld, M. E., Michael, S. I., Feng, M. Z. and Curiel, D. T. (1996) Targeted gene delivery by tropism-modified adenoviral vectors Nat. Biotechnol., 14, 1574-1578. Eastham, J. A., Chen, S. H., Sehgal, I., Yang, G., Timme, T. L., Hall, S. J., Woo, S. L. C. and Thompson, T. C. (1996) Prostate cancer gene therapy: Herpes simplex virus thymidine kinase gene transduction followed by ganciclovir in mouse and human prostate cancer models Hum. Gene Ther., 7, 515-523. Eastham, J. A., Hall, S. J., Sehgal, I., Wang, J. X., Timme, T. L., Yang, G., Connell-Crowley, L., Elledge, S. J., Zhang, W. W., Harper, J. W. and Thompson, T. C. (1995) In vivo gene therapy with p53 or p21 adenovirus for prostate cancer Cancer Res., 55, 5151-5155. Garlepp, M. J., Loh, S., Ramshaw, I. A., Leong, C. and Marley, J. V. (1997) Expression of B7-1 plus IL-2 after avipoxvirus transfer inhibits tumour development by a non-immunogenic murine mesothelioma cell line The Boden Conference on Gene Therapy, Thredbo, NSW. Golumbek, P. T., Hamzeh, F. M., Jaffee, E. M., Levitsky, H., Lietman, P. S. and Pardoll, D. M. (1993) Herpes simplex-1 virus thymidine kinase gene is unable to completely eliminate live, non-immunogenic tumor cell vaccines J. Immunotherapy, 12, 224-230. Gough, N., Gough, J., Metcalf, D., Kelso, A., Grail, D., Nicola, N., Burgess, A. and Dunn, A. (1984) Molecular cloning of cDNA encoding a murine haematopoietic growth regulator, granulocyte-macrophage colony stimulating factor Nature, 309, 763-7. Hall, S. J., Mutchnik, S. E., Chen, S. H., Woo, S. L. C. and Thompson, T. C. (1997) Adenovirus-mediated herpes simplex virus thymidine kinase gene and ganciclovir therapy leads to systemic activity against spontaneous and induced metastasis in an orthotopic mouse model of prostate cancer Int. J. Cancer, 70, 183-187. He, D. L., Mu, Z. M., Le, X. F., Hsieh, J. T., Pong, R. C., Chung, L. W. K. and Chang, K. S. (1997) Adenovirus-mediated expression of PML suppresses growth and tumorigenicity of prostate cancer cells Cancer Res., 57, 1868-1872. Hrouda, D. and Dalgleish, A. G. (1996) Gene therapy for prostate cancer Gene Therapy, 3, 845-852. Israeli, R., Grob, M. and Fair, W. (1997) Prostate-specific membrane antigen and other prostatic tumor markers on the horizon Urol. Clin. North Am., 24, 439-50. Israeli, R. S., Powell, C. T., Corr, J. G., Fair, W. R. and Heston, W. D. W. (1994) Expression of the Prostate-Specific membrane antigen Cancer Res ., 54, 1807-1811. Kawakita, M., Rao, G. S., Ritchey, J. K., Ornstein, D. K., Hudson, M. A., Tartaglia, J., Paoletti, E., Humphrey, P. A., Harmon, T. J. and Ratliff, T. L. (1997) Effect of canarypox virus (ALVAC)-mediated cytokine expression on murine prostate tumor growth J. Nat. Cancer Inst., 89, 428-436. Khatri, A., Xu, Z. Z. and Both, G. W. (1997) Gene Expression by Atypical Recombinant Ovine Adenovirus Vectors During Abortive Infection of Human and Animal Cells In Vitro Virology, Accepted for publication. Ko, S.-C., Gotoh, A., Thalmann, G. N., Zhau, H. E., Johnston, D. A., Zhang, W.-W., Kao, C. and Chung, L. W. K. (1996) Molecular therapy with recombinant p53 adenovirus in an androgen-independent, metastatic human prostate cancer model Hum. Gene Ther., 7, 1683-1691. Lee, C. H., Liu, M., Sie, K. L. and Lee, M. S. (1996) Prostate-specific antigen promoter driven gene therapy targeting DNA polymerase-alpha and topoisomerase II alpha in prostate cancer Anticancer Res., 16, 1805-1811. Lockett, L., Molloy, P., Russell, P. and Both, G. (1997) Relative efficiency of tumor cell killing in vitro by two enzyme prodrug systems delivered by identical adenovirus vectors Clin. Cancer Res., 3, 2075-2080. Moody, D., Robinson, J., Ewing, C., Lazenby, A. and Isaacs, W. (1994) Interleukin-2 transfected prostate cancer cells generate a local antitumor effect in vivo Prostate, 24, 244-51. Mueller, D., Jenkins, M. and Schwartz, R. (1989) Clonal expansion versus functional clonal inactivation: a costimulatory signalling pathway determines the outcome of T cell antigen receptor occupancy Annu. Rev. Immunol., 7, 445-80. Nandi, D., Gross, J. and Allison, J. (1994) CD28-mediated costimulation is necessary for optimal proliferation of murine NK cells J. Immunol., 152, 3361-9. O'Shea, J. and Ortaldo, J. R. (1992) The biology of natural killer cells: insights into the molecular basis of function Chap.1 IN "The Natural Immune System. The Natural Killer Cell", edited by C. Lewis and J. D. McGee (IRL Press at Oxford University Press, Oxford) pages 1-27. Pang, S., Dannull, J., Kaboo, R., Xie, Y. M., Tso, C. L., Michel, K., de Kernion, J. B. and Belldegrun, A. S. (1997) Identification of a positive regulatory element responsible for tissue-specific expression of prostate-specific antigen Cancer Res., 57, 495-499. Rodriguez, R., Schuur, E. R., Lim, H. Y., Henderson, G. A., Simons, J. W. and Henderson, D. R. (1997) Prostate attenuated replication competent adenovirus (ARCA) CN706: A selective cytotoxic for prostate-specific antigen-positive prostate cancer cells Cancer Res., 57, 2559-2563. Sanda, M. G., Ayyagari, S. R., Jaffee, E. M., Epstein, J. I., Clift, S. L., Cohen, L. K., Dranoff, G., Pardoll, D. M., Mulligan, R. C. and Simons, J. W. (1994) Demonstration of a rational strategy for human prostate cancer gene therapy J. Urol., 151, 622-628. Siders, W. M., Halloran, P. J. and Fenton, R. G. (1996) Transcriptional targeting of recombinant adenoviruses to human and murine melanoma cells Cancer Res., 56, 5638-5646. Silver, D., Pellicer, I., Fair, W., Heston, W. and Cordon-Cardo, C. (1997) Prostate-specific membrane antigen expression in normal and malignant human tissues Clin. Cancer Res., 3, 81-85. Smith, K. (1988) Interleukin-2: inception, impact, and implications Science, 240, 1169-76. Sorscher, E. J., Peng, S., Bebok, Z., Allan, P. W., Bennett, L. L., Jr and Parker, W. B. (1994) Tumor cell bystander killing in colonic carcinoma utilizing the Escherichia coli DeoD gene to generate toxic purines Gene Ther., 1, 233-238. Vesalainen, S., Lipponen, P., Talja, M. and Syrjanen, K. (1994) Histological grade, perineural infiltration, tumour-infiltrating lymphocytes and apoptosis as determinants of long-term prognosis in prostatic adenocarcinoma Eur. J. Cancer, 30A, 1797-803. Vieweg, J., Rosenthal, F. M., Bannerji, R., Heston, W. D. W., Fair, W. R., Gansbacher, B. and Gilboa, E. (1994) Immunotherapy of prostate cancer in the Dunning rat model - use of cytokine gene modified tumor vaccines Cancer Res., 54, 1760-1765. Vile, R. and Russell, S. J. (1994) Gene transfer technologies for the gene therapy of cancer Gene Therapy, 1, 88-98. Vrati, S., Macavoy, E. S., Xu, Z. Z., Smole, C., Boyle, D. B. and Both, G. W. (1996) Construction and transfection of ovine adenovirus genomic clones to rescue modified viruses Virology, 220, 200-203. Wickham, T. J., Mathias, P., Cheresh, D. A. and Nemerow, G. R. (1993) Integrin-alpha-v-beta-3 and integrin-alpha-v-beta-5 promote adenovirus internalization but not virus attachment Cell, 73, 309-319. Wright, G. J., Grob, B., Haley, C., Grossman, K., Newhall, K., Petrylak, D., Troyer, J., Konchuba, A., Schellhammer, P. and Moriarty, R. (1996) Upregulation of prostate-specific membrane antigen after androgen-deprivation therapy Urology, 48, 326-34. Wu, T., Huang, A., Jaffee, E., Levitsky, H. and Pardoll, D. (1995) A reassessment of the role of B7-1 expression in tumor rejection J. Exp. Med., 182, 1415-21. Xu, Z. Z., Hyatt, A., Boyle, D. B. and Both, G. W. (1997) Construction of ovine adenovirus recombinants by gene insertion or deletion of related terminal region sequences Virology, 230, 62-71. Acknowledgements The work was supported by grants from the National Health and Medical Research Council, Australia (1995-1997) and from the Cancer Council, New South Wales (1998-1999). Copyright 1998 Australian Biotechnology Association Ltd. The following images related to this document are available:Line drawing images[au98019b.gif] [au98019d.gif] [au98019c.gif] [au98019a.gif] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}