 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Volume 8 Number 3, May/June 1998, pp. 148-152 Adjuvants John C Cox, R&D Division, CSL Limited, 45 Poplar Road, Parkville, Victoria 3052 Australia
Code Number:AU98022
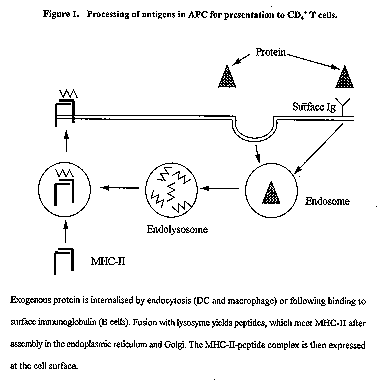
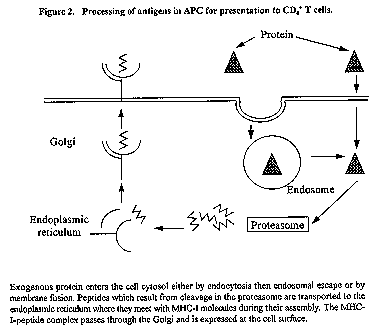
Adjuvants can act in a number of diverse ways to help the immune system respond to a vaccine. An understanding of these modes of action along with some knowledge of the epidemiology, pathogenesis and the required protective response to the disease of relevance can permit selection of an adjuvant or adjuvant combination which will give maximum benefit. This review will consider these modes of adjuvant action and discuss issues of formulation which are important to adjuvant function. Introduction Adjuvants (from the Latin adjuvare = to help) have been used to improve vaccine efficacy from the early 1920s and since that time, the scientific and patent literature has been increasingly bombarded with articles on various substances and formulations with purported adjuvant activity. Generally these were uses with "traditional" immunogen-based vaccines eg. bacterial toxoids, although recently, adjuvants which help DNA vaccines have also been described. The reader is referred to two earlier reviews (Cox & Coulter 1992; 1997) for their extensive descriptions and bibliography. This review will present an overview of these papers along with discussion of new concepts and applications of adjuvants. How do adjuvants act? We have proposed (Cox & Coulter, 1997) that adjuvants can act in one or more of five different ways. One of these, immunomodulation, requires no association or interaction between adjuvant and immunogen, and in fact immunomodulatory adjuvants can generally be effective when administered at a separate time or site to the immunogen. Conversely, the other four modes of adjuvant action benefit or perhaps even depend upon a level of association between adjuvant and immunogen which will persist in vivo from several hours in the case of presentation, targeting and CTL induction through to weeks and even months in the case of depot formation. Immunomodulation Immunomodulation generally involves an up-regulation of the cytokine network. This may be a general up-regulation, where the magnitude of the immune response is increased without altering the nature of the response, or alternatively it may be selective with an up-regulation of one T-helper subset and a concomitant down-regulation of the other. Antigen presentation Antigen presentation describes the ability to present antigen in conserved native conformation, preferably in multimeric form. At least three series of events are required to achieve an effective antibody response and these are summarised in Figure 1. Firstly, antigen is endocytosed by antigen presenting cells (APC) typically either dendritic cells (DC) or less importantly macrophages, and digested within the endolysosome to peptides and other fragments. Peptides, possibly randomly selected, associate with already formed major histocompatibility complex class II (MHC-II) antigen and pass to the surface of the APC. In the second series of interactions, the fully mature APC, having migrated from its peripheral site to a lymphoid organ, presents its MHC-II - peptide complex to CD4+ T helper cells. Those with complementary T cell receptors specific to this complex, in the presence of appropriate co-stimulatory signals, undergo clonal expansion and/or differentiation. Figure 1. Processing of antigens in APC for presentation to CD4+ T cells. The third set of interactions involves B cells. These also are APCs but with the specialist function of producing antibody. All B cells have imunoglobulin (Ig) on their surface. Each B cell will have Ig of a single specificity but within the body every possible Ig variant will be on the surface of some B cells. This Ig will recognise and bind to specific antigenic determinants whereupon the antigen will be endocytosed and processed just as was previously described for DCs. These B cells will now attract CD4+ T cells which have already been clonally expanded by interaction with DCs in stage 2 and these T cells will now help the B cell to clonally expand and differentiate to plasma cells which will produce antibody of the same specificity as that on the surface of their parent B cell. The important message from this is that if denatured antigen is presented to B cells, the same overall process will occur but the result will be antibody with little if any protective ability i.e. it will not recognise and neutralise native toxins or antigens on the surface of a pathogen. The importance of antigenic presentation is to maximise the production of useful neutralising antibody. CTL induction CTL induction requires processing of antigen through the class 1 or cytosolic pathway (see Figure 2) which involves digestion within proteasomes, a 26S specialised proteolytic enzyme complex, then incorporation of peptides of defined length (8 to 10 amino acids) into the closed groove of an MHC-I molecule during its assembly in the endoplasmic reticulum. This complex is then presented on the surface of cells and attracts CD8+ T cells with complementary receptors. Again, in the presence of appropriate co-stimulatory signals, these CD8+ cells will expand into an army of CTL cells with specificity to cells bearing the specific MHC-I - peptide complex. Whereas only APCs (DCs, macrophages and B cells) can produce MHC-II, nearly all cells in the body can produce MHC-I which, when surface-exposed in association with foreign peptide, is the body's mechanism to indicate "foreigness" due to any aberrant intracellular event, and acts as a request for elimination. It is currently a moot point whether all cells or just APCs can induce CTL responses; certainly APCs are most efficient. From an adjuvant point of view, CTL induction requires delivery of an antigen to a cell compartment (the cytosol) which is not the normal endosomal process of uptake and processing. The most likely ways for adjuvants to achieve this is by fusion with cell membranes (the probable mode of action of liposomes) or by endosomal disruption (the probable mode of action of saponins and other surface-active adjuvants). In either case, association between antigen and adjuvant is an important requirement for this action, and therefore must be considered during vaccine formulation. Figure 2. Processing of antigens in APC for presentation to CD4+ T cells Targeting Targeting describes mechanisms whereby adjuvants can help deliver antigen to APC and hence reduce the amount of antigen required in a vaccine formulation for a desired magnitude of immune response. This can be achieved in a number of ways including targeting of sugar residues on adjuvants to cell surface receptors (eg. mannan to mannose receptors on APCs), targeting of proteins to cell surface sugars (eg. cholera toxin (CT) to GM-1 ganglioside) and the formation of small particles which are more readily endocytosed (eg. liposomes, Iscoms). Depot formation Depot formation may be either short-term or long-term. Short-term depots slowly release antigen to the immune system over one to two weeks, and are typified by water-in-oil emulsions (eg. Freund's adjuvant) and aluminium salts. Surgical removal of the dose site within 1 week of inoculation usually reduces the magnitude of the response but removal after 2 weeks has no effect. Long-term depots are designed to hold the vaccine components at the dose site without release for weeks or even months and then to make them available to APCs, preferably over a short period i.e. as a pulse. In this way a two or three course vaccine can be given as a single dose. Options for adjuvants to modify immune responses From the previous section it should be clear that adjuvants can modify both the magnitude and duration of an immune response and the quality or type of response. In selecting an adjuvant for a vaccine it is of paramount importance to know firstly what type of immune response is likely to be efficacious. In the absence of this knowledge, immune responses may be generated which are detrimental to protection and recovery. Once the correct type of immune response has been chosen, then its magnitude and duration can be improved by the correct selection of adjuvant combinations. Adjuvants are able to induce three broadly-different types of immune response: Induction of Th2 responses These responses are associated with IL-4 and IL-5 cytokines and generally result in production by plasma cells of non-complement fixing antibodies (IgG1 in mice, IgG2 in humans, sIgA in all species). These are non-inflammatory antibodies which can bind to microbes or microbial products (eg. toxins) and prevent their harmful effect. The resulting neutralised complexes are rapidly removed by macrophages or NK cells. Induction of secretory IgA (sIgA) seems to require activation of mucosal lymphoid tissue; sIgA, the most effective mucosal antibody, owes its effectiveness to its resistance to proteases and massive in vivo production. Induction of IgE, which also appears to be an outcome of a Th2 response requires IL-13 for its induction and has both good effects (it is important in metazoan killing) and bad effects (associated with allergic reactions). Immunomodulatory adjuvants are required to maximise Th2 responses. Aluminium salts (alum, aluminium phosphate, aluminium hydroxide) are strong Th2 inducers but unfortunately also induce IgE. Other Th2 inducers include vitamin D3, water soluble muramyl dipeptide (MDP) analogues, poly A:U and CT (cholera taxin) or LT (labile toxin from E.coli). Induction of Th1 responses These responses are associated with the cytokines IL-2, IL-12 and gIFN and generally result in production by plasma cells of complement fixing antibodies (IgG2a in mouse, IgG1 and IgG3 in man). These antibodies will bind to their targets then activate complement by the "classical pathway" starting with C1q. Macrophages become far more activated in the presence of both antibody and complement and kill surrounding pathogens by a range of mediators including reactive oxygen intermediates and nitric oxide. Mechanisms fortunately exist to minimise killing of autologous cells. Different immunomodulatory adjuvants are required to maximise Th1 responses and include lipid A and analogues (the adjuvant active region of bacterial endotoxin), DHEA (dehydro epiandrosterone, a steroid analogue), TDM (trehalose dimycolate, a component of mycobacterial cell walls) and poly I:C. Generally, induction of Th1 responses has a negative feedback on Th2 responses and vice versa. However, saponin adjuvants (QuilA, QS21, Iscoms) increase the magnitude of both Th1 and Th2 responses. Induction of CTL responses Frequently, microbes can grow within cells without expressing foreign proteins on the cell surface, and similarly tumorigenic changes within a cell can occur without foreign surface antigens being expressed. However, in most cases there are foreign intracellular proteins which are available for processing within the cell cytosol. An outcome of this processing is presentation by these cells of foreign peptide - MHC-I complexes on their surface. These cells are a target for CD8+ cytotoxic T lymphocytes which, when activated, can kill large numbers of autologous cells bearing foreign peptide by injection of granzymes and perforins, which result in cell lysis. Adjuvants which induce a CTL response must be able to deliver antigen (or peptide) to APC in such a way that peptides can be expressed in association with MHC-I molecules. In practical terms, this will involve close association between adjuvant and antigen in vivo. Saponin adjuvants are the most effective way to induce CTL responses and probably act by causing endosomal escape of antigen. Liposomes are also effective, probably by cell membrane fusion where associated antigen is transferred directly into the cytosol. In specific cases, water-in-oil emulsions can deliver peptides directly to empty MHC-I on the surface of an APC and, in the presence of a signal (eg. a foreign protein in the formulation), cause the APC to mature, resulting in CTL induction. Matching immune responses with target diseases Prophylactic vaccines are designed to prevent infection or, more realistically, limit infection to a sufficiently low level that clinical symptoms are minimal or absent. This can generally be achieved with neutralising antibody hence the preferred adjuvant will induce high and prolonged titres of antibody which can recognise native epitopes on the surface of the pathogen. Frequently a Th2 response may be preferred because this can lead to sIgA and a minimal inflammatory response at the site of invasion (usually a mucosal surface). Experimentally (in mice) mucosal responses are best generated with CT, or LT but in man these are unacceptably toxic. Various non-toxic mutants are currently under evaluation for human oral use; alternatively CTB or LTB coupled to antigen may be effective for intranasal delivery. Intranasal delivery of Iscoms has also been shown to induce good sIgA levels and protection to challenge in mice, and has the added advantage of excellent antigen presentation in addition to the desired immunomodulatory activities. Although induction of sIgA may seem desirable, there is considerable practical evidence that vaccines adjuvanted with aluminium salts and administered parenterally can protect against mucosal pathogens (eg. B. pertussis, H. influenzae) and influenza vaccines delivered parentally without adjuvant have good efficacy. This is probably because any lesion caused by an invading pathogen at a mucosal surface will immediately expose that pathogen to antibodies in the systemic circulation. There are many pathogens where, once initial colonisation has occurred, antibody alone is unable to resolve (HSV) or even limit (HIV, M.tuberculosis) the infection. In these situations, some level of cellular immunity is essential. This may be predominantly Th1 induced (eg. M.tuberculosis, HSV?) or alternatively may require effective CTL responses (HIV, HCV). Because there seems to be an inexorable trend for the immune system to progress from a Th1 to a Th2 response, it is important with these vaccines to use an adjuvant which can establish and maintain a Th1 response. Similarly, many viruses are able to mutate in vivo to modify peptide sequences which can act as CTL targets. To counter this, a vaccine should stimulate CTL responses to a broad range of targets so that sterilising immunity can be achieved before the pathogen has been able to mutate sufficiently. Therapeutic vaccines are designed to either terminate latent or persistent infections or to eliminate cancer cells. In either case, it is clear that the immune response has been unable to deal with the problem despite a lengthy exposure. There are many reasons why this may be so, including down-regulation of MHC-I expression in infected cells, deletion of potential target sequences from the genome, lack of exposure of any foreign protein or sequence to the immune system and inability of the affected cell to induce a CTL response. Lack of antibody is usually not the problem although in the case of H. pylori infection, a Th1 rather than Th2 antibody response may be the cause of inflammation and hence continued colonisation. However, generally it is likely that CTL and/or other cellular responses will be required in immunotherapy, thus the selected adjuvant will need to induce Th1 and CTL. What adjuvants can be used in humans? Virtually all adjuvanted human vaccines use aluminium salts as adjuvant. Because these adjuvants are both immunomodulatory and able to form short-term depots, it is important that antigen be sufficiently strongly associated with the adjuvant to persist for several weeks in vivo in the depot. Binding is by electrostatic attraction, and because aluminium phosphate and aluminium hydroxide have opposite charges at pH7.0 it is likely that one or other will bind most antigens. Calcium phosphate is a rarely used vaccine adjuvant licenced in France for use in humans. Water-in-oil (w/o) adjuvants (as Freund's Incomplete adjuvant) were used extensively in human vaccines in the 1950s but their use was discontinued because of a low but unacceptable rate of dose-site reactions. Squalene-based w/o adjuvants (Seppic) are currently in human trial. Oil-in-water (o/w) vaccines have received considerable attention and one (MF59, Chiron) is the basis of a registered influenza vaccine in Italy. Other o/w vaccine formulations incorporate various immunomodulators and are currently in clinical trial. Of these, the most important are the SB formulation which contains the saponin QS21 and MPL a lipid A derivative, and the Ribi formulation which contains MPL and CWS, a bacterial cell wall skeleton. These formulations contain biodegradable oil and are in phase III clinical trial. Saponin-based adjuvants are in clinical trial either as Iscoms or as QS21 either alone or in an o/w formulation. Liposomes containing lipid A are also receiving attention in the clinic. There have not yet been serious studies of microencapsulated vaccines in humans, but there seems to be no major impediment to their use and they offer the opportunity to deliver a full vaccine course in a single dose, an option attractive to developed countries and of major importance in developing countries. Summary A rational choice of adjuvant (or adjuvant combination) needs a knowledge of the type of immune response required for protection along with an understanding of the modes of action of various adjuvants in the target species. It is likely that any effective vaccine adjuvant will combine an immunomodulatory adjuvant into a formulation which will permit association of immunogen and adjuvant so that the other modes of adjuvant action can be fully realised. In many cases a choice will need to be made. For example, presentation and short-term depots are usually not mutually achievable because of the need to sequester antigen within the depot rather than expose it at the surface. In other situations, a clear choice must be made between Th1 and Th2. The need for CTL responses should be carefully considered because, if important, the number of adjuvant options is limited which in turn can restrict choices for other modes of adjuvant action. Thus, if Iscoms are chosen because of a wish to induce CTL responses, the vaccine will also acquire the immunomodulatory activities of saponins. Finally, although microencapsulation is not compatible with oil-based adjuvants, new techniques in microencapsulation permit many other adjuvants to be incorporated within the microcapsule thus their benefits can be superimposed on the benefits of the delayed-release formulation. Bibliography Cox, JC and Coulter, AR. Advances in adjuvant technology and application. In: Animal Parasite Control Utilizing Biotechnology (Ed. Yong WK). CRC Press. Boca Raton, 1992, P.49-112. Cox, JC and Coulter, AR. Adjuvants - a classification and review of their modes of action. Vaccine 1997, 15, 248-256. Eun, HM and Aubert, A. Categories of Vaccines according to their antigenic target: (Anti-) viral vaccines In: Veterinary Vaccinology (Eds. Pastoret, PP; Blancou, J; Vannier, P and Verschueren, C) Elsevier, Amsterdam, 1997 p 435-448. Roitt, I. Essential Immunology. Blackwell Scientific Publications, Oxford 1994. Vogel, FR and Powell, MF. A compendium of vaccine adjuvants and excipients. In:
Vaccine
Design, The Subunit and Adjuvant Approach (Eds. Powell, MF and Newman, MJ) Plenum
Press New York 1995, p. 141-228. Copyright 1998 Australian Biotechnology Association Ltd. The following images related to this document are available:Line drawing images[au98022b.gif] [au98022a.gif] |
| |||||||||
{kind=link}
{kind=link}