 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Volume 8 Number 3, May/June 1998, pp. 153-159 DNA vaccines: Will they deliver? Boyle JS1, Agius CT2, Lew AM1 and Barr IG 1Co-operative Research Centre for Vaccine Technology, Walter and Elisa
Hall Institute of
Medical Research, P.O. Box RMH, Melbourne, Australia, 3050. email: boyle_j@wehi.edu.au
Code Number:AU98023
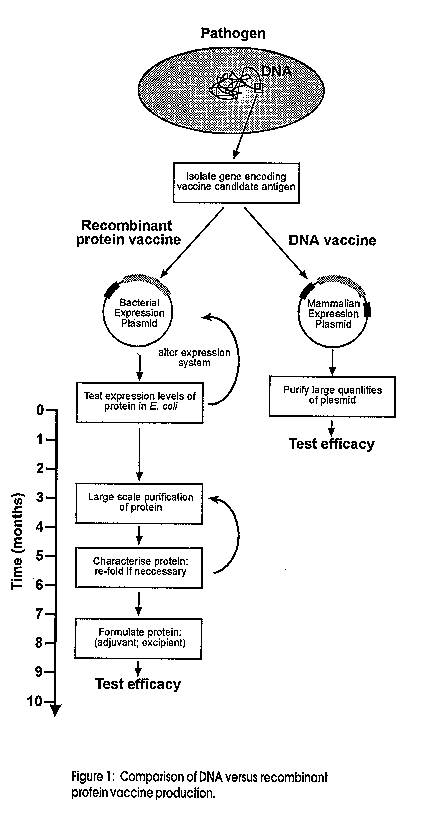
A new form of vaccination using DNA as the vaccine instead of protein or attenuated pathogens has the potential to revolutionise the way vaccines are made and used. Based on administering purified plasmid DNA, these vaccines have shown remarkable promise in numerous animal models for the prevention of several important human diseases. The use of DNA as a vaccine raises several questions including the basic questions of just how do they work to stimulate an immune response, many of which we are only beginning to understand. In addition to efficacy, there are also several safety issues that will have to be addressed prior to large scale testing in humans. Given the excitement surrounding this new technology, the overriding question is whether they will deliver some or indeed any of their potential benefits to mankind. Introduction The possibility of vaccinating effectively with DNA has ignited vaccine research in the 1990's. From the first observations that a DNA vaccine could protect laboratory animals from influenza infection (Ulmer et al. 1993) DNA vaccines have rapidly progressed and are already being tested in a number of human clinical trials. These and subsequent trials will establish whether this first generation of DNA vaccines will be safe and useful for the prevention and treatment of human diseases. This review will give a brief introduction to DNA vaccines and will outline some of their promises and problems. What are DNA vaccines? DNA vaccines differ from conventional vaccines, which can be obtained from whole killed or attenuated organisms or more recently proteins made by recombinant techniques. DNA vaccines consist of a plasmid encoding a vaccine antigen as well as other sequences that enable the expression of the protein in mammalian cells. The foreign protein (selected from the organism e.g. Hepatitis B surface antigen) is then synthesised by the recipient's own cells and an immune response to the encoded protein is initiated. Consequently there is no need to grow the organism, inactivate or attenuate it nor purify or express recombinant components. DNA is also physicochemically very stable. Thus DNA vaccines have the potential to simplify dramatically the production, purification, storage and combination of different vaccines. This could save enormous amounts of time and investment in evaluating vaccine candidate antigens in the laboratory and in producing vaccines (Figure 1). However, it is imperative that DNA vaccines result in a sustained and protective immune response. There is a rapidly growing body of evidence that many proteins from a range of infectious organisms (viral, bacterial and parasitic) can be given as DNA vaccines and confer protection in numerous animal species (Donnelly et al. 1994). A list of the principal characteristics that DNA vaccines offer is given in Table 1. Figure 1. Comparison of DNA versus recombinant protein vaccine production Table 1: Characteristics of DNA vaccines
Why all the excitement? While vaccination has enjoyed a number of stunning successes such as the elimination of smallpox and near eradication of polio and despite much work, only some 25 vaccines against human pathogens are currently available. There is a desperate need for at least this number of new vaccines against emerging human diseases. For example, in the developed world, vaccines are needed to combat Hepatitis C, D, and E, HIV-1 and tuberculosis, while in the developing world, Malaria, Leishmania and many other diseases still persist. Astonishingly, in the last few years a number of these diseases have been prevented in experimental animal models with the use of DNA vaccines. Furthermore, DNA vaccines may have some potential in treating cancer, autoimmune disease and asthma due to their ability to induce cytotoxic T cells or to modify ongoing immune responses. DNA immunisation: mechanisms of immune induction At present there is a limited understanding of the mechanisms underlying the induction of immune responses following DNA immunisation. It is known that both antibody and cellular (cytotoxic T cell) responses are induced following DNA immunisation. This is despite the very small amounts of antigen being produced (in the order of nanograms) (Wolff et al. 1990). however this expression is continuous (Wolff et al. 1991). Antibody responses cannot be detected when such small amounts of soluble antigen are injected as a bolus, which raises questions as to why and indeed how antibody induction occurs after DNA vaccination. The studies into antigen expression following intramuscular (i.m.) DNA immunisation have shown that myocytes are the primary targets for transfection. Myocytes (both myoblasts and myotubes) express MHC Class I molecules, for presentation of antigen to CD8 + T cells, at low levels but not MHC Class II, for presentation to CD4+ T cells. Both Class I and Class II expression is increased after exposure to the cytokine INF-g, with Class II only detected in 40 to 90% of cells (Hohlfeld andEngel 1994). However, myocytes lack expression of other molecules such as the co-stimulator B7 that is necessary for efficient immune induction (Lenschow et al. 1992; Linsley et al. 1992). Therefore the role of the transfected myocyte in immune induction is unclear. On the other hand, intradermal (i.d.) injection of DNA may directly transfect antigen presenting cells, which are potent initiators of immune responses (Raz et al. 1994). Mechanism of CTL induction CTLs are important in the clearance of virally infected cells and may be crucial for the clearance of cancerous cells. They may also be involved in infections with some intracellular bacteria such as Mycobacteria (Lowrie 1998). The ability to induce CTL responses with DNA immunisation is a major advantage for vaccination purposes because there are very few methods by which CTL responses can be reliably induced. These include injection of wild type or attenuated virus containing antigen-encoding sequences, liposome preparations, use of Freund's complete adjuvant and transfer of in vitro transfected (Manickan et al. 1997) or osmotically loaded autologous cells (Moore et al. 1988). In general, exogenous soluble antigen in the absence of any adjuvant does not prime a CTL response very efficiently. There are several possibilities that could account for CTL induction following DNA immunisation (Pardoll and Beckerleg 1995). Firstly, the transfected myocyte may present endogenously produced antigen on Class I and prime a CTL response directly. This is unlikely because of the lack of expression of essential accessory molecules such as B7 on myocytes. Secondly, an antigen could be transferred to an antigen presenting cell, which then primes the CTL response, a mechanism known as cross-priming. Finally, antigen presenting cells which prime a CTL response could be directly transfected. In the case of i.d. DNA immunisation the latter mechanism may play a role because cells with a dendritic morphology have been shown to express encoded proteins after DNA injection (Raz, et al. 1994; Condon et al. 1996). Therefore, if different cell types are transfected after i.m. or i.d. DNA injection, it is possible that the mechanisms underlying the induction of CTL responses also differ. Recently, there have been studies directly examining the mechanism of CTL induction after i.m. DNA immunisation (Corr et al. 1996; Doe et al. 1996; Ulmer et al. 1996). Together these studies provide strong evidence that priming of CTL responses after i.m. DNA immunisation occurs via cross-presentation of antigen by non-muscle cells of bone marrow origin. It is unclear if this cross-priming mechanism is responsible for priming CTL responses when DNA is injected or propelled with a gene gun into other sites but there is some evidence that cross-priming may also occur after biolistic transfection (Iwasaki et al. 1997). Mechanism of antibody induction It remains puzzling as to how such small amounts of antigen can induce antibody responses after DNA immunisation. The continuity of exposure or accumulation of antigen may be critical. It has been shown that for bolus administration of antigen, relatively high doses are required to induce an antibody response (Petty et al. 1972) whereas antibody responses, albeit in only 3 of 10, mice can be induced after daily administration of small doses (Petty and Steward 1976). Interestingly, it was shown that the kinetics of the antibody response to hepatitis B antigens peaked at 4 to 8 weeks after DNA immunisation, which closely mimicked natural viral infection (Michel et al. 1995). The IgG subclass response to a herpes simplex virus antigen (gB) after DNA immunisation of mice was examined and shown to be biased towards an IgG2a dominance as was the case after infection with wild type virus (Manickan et al. 1995). Subsequently, others have reported an IgG2a dominance in the antibody response after DNA immunisation in mice (Cardoso et al. 1996; Pertmer et al. 1996; Raz et al. 1996). At least for i.d. DNA immunisation, it was suggested that the DNA itself could account for the induction of antibody responses to such low levels of antigen via immunostimulatory sequences present in the bacterial DNA (Sato et al. 1996). It had been shown that unmethylated CpG motifs in bacterial DNA can stimulate B cells (Krieg et al. 1995) and T cells to proliferate and produce cytokines such as INF-g (Klinman et al. 1996; Yi et al. 1996) and activate antigen presenting cells (Stacey et al. 1996). It was suggested later that the immunostimulatory sequences were responsible for the apparent IgG2a bias in the antibody response after i.d. DNA immunisation (Roman et al. 1997) via stimulation of CD4 T cells to produce cytokines (INF-g, INF-a, INF-b and IL-12). Furthermore, co-injection of oligonucleotides containing a CpG motif increased IgG2a levels to a soluble exogenous antigen. However, other factors including route of immunisation (Mor et al. 1995), form of antigen (Cardoso et al. 1996) and choice of promoter may also be important in determining the level and type of antibody produced. Safety concerns with DNA vaccines Besides efficacy, safety concerns for DNA vaccines must be addressed if this technology is ever to be introduced into the clinic. There are several safety concerns regarding DNA vaccines that have already been raised and these are being addressed in ongoing animal testing and human trials as discussed below. There is a possibility that integration of the DNA vaccine plasmid into the host cell genome either randomly or by homologous recombination could lead to insertional mutagenesis. Such an event might result in activation of oncogenes, inactivation of tumour suppressor genes or chromosomal instability. Thus far, experimental evidence has shown that the risk of integration is extremely low. In a study by Nichols et al. (1995), no integration could be detected to a sensitivity of 1-7.5 plasmid copies per 150,000 nuclei after injecting mice intramuscularly with 200ug of plasmid DNA. At this level of detection, the mutation frequency induced by DNA integration would be approximately three orders of magnitude less than the natural frequency. Another study attempted to detect integration events by sequencing regions of chromosomal DNA obtained from the site of DNA injection. No evidence of plasmid/chromosome junctions (indicating integration) was observed although only 1800 samples were examined (Wolff et al. 1992). DNA vaccines may alter the immune state, resulting in tolerance or autoimmunity such as the induction of anti-DNA antibodies. Repeated injection of small quantities of antigen can lead to the development of immunologic unresponsiveness. It is thought that the persistent expression of small amounts of antigen produced by DNA vaccination might reduce responses after a pathogenic challenge rather than protection. In neonatal mice, a DNA vaccine encoding the circumsporozoite protein of the malaria parasite has been shown to induce tolerance (Mor et al. 1996). In contrast, DNA vaccination was found to protect newborn chimpanzees against a challenge infection with hepatitis B virus (Prince et al. 1997). In adult mice, DNA vaccines have not induced tolerance even after subimmunogenic doses (Liu et al. 1997). Another concern is that DNA vaccine may lead to autoimmunity whereby the muscle cells producing antigen become targets for immune attack or the DNA induces self-reactive anti-DNA antibodies (as occur in systemic lupus erythematosus). One study directly addressed these concerns and could not demonstrate vaccination leading to autoimmunity even in genetically susceptible strains of mice (Mor et al. 1997). Together these studies suggest that DNA vaccination would result in the generation of an immune response rather than tolerance without inducing autoimmunity. The possibility of systemic toxic effects or local site reactivity due to contaminants in the DNA preparations has also been raised. One such contaminant is endotoxin from E. coli, which can co-purify with plasmid DNA. It has been shown that the presence of endotoxin in a DNA vaccine caused dose-related toxicity, increased serum IgM levels (Wicks et al. 1995) and in certain circumstances reduced immune responses (Boyle et al. 1998a) in mice injected with the vaccine. Endotoxin can be readily removed down to trace levels with the use of commercial reagents or using triton X-114 to extract the endotoxin (Aida and Pabst 1990). The inherent stability and charge of DNA as compared to most proteins should be of great benefit for enabling the easy removal of other contaminants from preparations if and when they are identified in the future. Regulatory hurdles for DNA vaccines In the USA, the introduction of DNA vaccines for human use is regulated by the Center for Biologics Evaluation and Research (CBER) which is part of the FDA. In October 1996, CBER issued a preliminary guideline for the production and quality control of DNA vaccines entitled "Points to Consider on Plasmid DNA Vaccines for Preventive Infectious Disease Indications". This document outlines the regulatory concerns associated with the manufacture, preclinical evaluation and clinical studies of DNA vaccines (a copy is available online at http://www.genweb.c om/Dnavax/Announce/plasmidptc.html). This document also reflects the safety concerns associated with the application of a new and largely uncharacterised system into clinical use. All new DNA vaccines must be fully characterised as to the vector components and their production with mandatory monitoring for contaminants such as RNA, protein and bacterial genomic DNA. Of note, CBER advised manufacturers against the use of b-lactam antibiotics (e.g. ampicillin) in vaccine constructs as these antibiotics can result in allergic reactions, instead recommending the use of aminoglycoside antibiotics such as kanamycin or neomycin. Furthermore, to limit the possibility of chromosomal integration, CBER recommended that plasmid DNA with strong homology to known sequences in the human genome should be avoided. Although each new DNA vaccine will be reviewed on a case by case basis, this CBER document does provide a starting point from which manufacturers can begin. Of course the safety of healthy subjects in clinical trials is paramount and pre-clinical data is reviewed carefully prior to commencement of trials. It is possible however, that DNA vaccines may be used for therapeutic intervention in life-threatening situations where the potential benefit is worth the hypothetical risk. Current human clinical trials with DNA vaccines Table 2: Current human DNA vaccine clinical trials
*These are not DNA vaccine trials but utilise essentially the same technology. Table 2 lists a number of human clinical trials completed or in progress with DNA vaccines. These are mainly Phase I trials to assess safety and have been undertaken mainly by commercial companies drawn by the success of DNA vaccines in animal models. Initial trials have investigated relatively low doses of plasmid DNA in the range of tens to a few hundred micrograms. The majority of these trials have investigated the intramuscular route of inoculation. However, one trial (PowderJect) have investigated the use of the gene gun - a biolistic delivery system that utilises helium to propel gold particles coated with the DNA vaccine directly into the skin of the patient. All of the current DNA vaccines in clinical trials have been given to healthy volunteers although some HIV-1 positive patients have been involved in some trials. Despite a number of these trials having been completed, no detailed clinical data has been published to date. Interestingly cancer treatments utilising the same vector as is used in DNA vaccines but carrying a cytokine gene (IL-2), or HLA-B7 instead of a vaccine gene have recently progressed to Phase I/II and Phase II trials respectively (Genetic News 1998 18, 1; 28). These vectors differ from other DNA vaccines in that they do not raise an immune response to the encoded proteins (IL-2 or HLA-B7) but rather will hopefully deliver immuno-modulatory signals. A press release (Washington D.C., April 30 1998) from the Hoffman trial stated that their malaria DNA vaccine was "well tolerated and safe, and that preliminary analysis of specimens from trial participants suggested a good cellular response with features they believe may be important in preventing disease". The research community keenly awaits the publication of the full results from all of these trials. How do I find out more information? Robert Whalen, one of the pioneers in the DNA vaccine field has established an excellent web site at http://www.genweb.com/Dnavax/dn avax.html. It contains the latest news as well as a complete reference database, patent information, protocols and a contacts list for people working on DNA vaccines. For more extensive reviews the reader is directed to a recent review by Donnelly et al. (1997). Conclusion Despite the remarkable results obtained with DNA vaccines in experimental laboratory animals the jury is still out on the ultimate applications (if any) for which DNA vaccines will provide in humans. It is unlikely however that DNA vaccines will replace existing vaccines. A more likely use for DNA vaccines is in preventing diseases for which there is significant morbidity or mortality associated with infection and where there is no vaccine or effective treatment available such as drug-resistant Tuberculosis, Hepatitis C and HIV-1. There is also the potential for the treatment of cancers where DNA vaccine induced CTL responses may play an important role in the control or elimination of tumours. It is also possible that this current generation of DNA vaccines will fail to generate the same responses in humans that has been achieved in murine models as has been suggested by relatively poor responses in larger animals such as cattle and sheep. There may need to be enhancements made to DNA vaccines in order to make them more effective in humans. Improvements to DNA vaccines have been shown by targeting antigen to sites of immune induction (Boyle et al. 1998b) or by the inclusion of plasmids expressing cytokine or co-stimulatory genes (Xiang and Ertl 1995). The next phase of DNA vaccine research awaits the release of the human clinical trial data to determine if in fact they have already delivered a useful means of disease prevention and treatment or a quantum leap in immunogenicity will be required for human applications. References Aida, Y. & Pabst, M.J. (1990) Removal of endotoxin from protein solutions by phase separation using Triton X-114 J. Immunol. Methods 132, 191-195. Boyle, J.S., Brady, J.L., Koniaras, C. & Lew, A.M. (1998a) The inhibitory effect of lipopolysaccharide on the immune response after DNA immunization is route dependent DNA Cell Biol. (in press). Boyle, J.S., Brady, J.L. & Lew, A.M. (1998b) Enhanced responses to a DNA vaccine encoding a fusion antigen that is directed to sites of immune induction Nature 392, 408-411. Cardoso, A.I., Blixenkrone-Moller, M., Fayolle, J., Liu, M., Buckland, R. & Wild, T.F. (1996) Immunization with plasmid DNA encoding for the measles virus hemagglutinin and nucleoprotein leads to humoral and cell-mediated immunity Virol. 225, 293-299. Condon, C., Watkins, S.C., Celluzzi, C.M., Thompson, K. & Falo, L.D., Jr. (1996) DNA-based immunization by in vivo transfection of dendritic cells Nature Med. 2, 1122-1128. Corr, M., Lee, D.J., Carson, D.A. & Tighe, H. (1996) Gene vaccination with naked plasmid DNA: mechanism of CTL priming J. Exp. Med. 184, 1555-1560. Doe, B., Selby, M., Barnett, S., Baenziger, J. & Walker, C.M. (1996) Induction of cytotoxic T lymphocytes by intramuscular immunization with plasmid DNA is facilitated by bone marrow derived cells Proc. Natl. Acad. Sci. USA 93, 8578-8681. Donnelly, J.J., Ulmer, J.B. & Liu, M.A. (1994) Immunization with DNA J. Immunol. Methods 176, 145-152. Donnelly, J.J., Ulmer, J.B., Shiver, J.W. & Liu, M.A. (1997) DNA Vaccines Ann. Rev. Immunol. 15, 617-648. Hohlfeld, R. & Engel, A.G. (1994) The immunobiology of muscle Immunol. Today 15, 269-274. Iwasaki, A., Torres, C.A., Ohashi, P.S., Robinson, H.L. & Barber, B.H. (1997) The dominant role of bone marrow-derived cells in CTL induction following plasmid DNA immunization at different sites J. Immunol. 159, 11-14. Klinman, D.M., Yi, A.K., Beaucage, S.L., Conover, J. & Krieg, A.M. (1996) CpG motifs present in bacteria DNA rapidly induce lymphocytes to secrete interleukin 6, interleukin 12, and interferon gamma Proc Natl Acad Sci U S A 93, 2879-2883. Krieg, A.M., Yi, A.K., Matson, S., Waldschmidt, T.J., Bishop, G.A., Teasdale, R., Koretzky, G.A. & Klinman, D.M. (1995) CpG motifs in bacterial DNA trigger direct B-cell activation Nature 374, 546-549. Lenschow, D.J., Zeng, Y., Thistlethwaite, J.R., Montag, A., Brady, W., Gibson, M.G., Linsley, P.S. & Bluestone, J.A. (1992) Long-term survival of xenogeneic pancreatic islet grafts induced by CTLA4lg Science 257, 789-792. Linsley, P., Wallace, P., Johnson, J., Gibson, M., Greene, J., Ledbetter, J., Singh, C. & Tepper, M. (1992) Immunosuppression in vivo by a soluble form of the CTLA-4 T cell activation molecule Science 257, 792-795. Liu, M.A., W., M., Ulmer, J.B., Shiver, J. & J., D. (1997) Immunisation of non-human primates with DNA vaccines Vaccine 15, 909-912. Lowrie, D.B. (1998) DNA vaccination exploits normal biology Nature Med. 4, 147-148. Manickan, E., Kanangat, S., Rouse, R.J., Yu, Z. & Rouse, B.T. (1997) Enhancement of immune response to naked DNA vaccine by immunization with transfected dendritic cells J Leukoc. Biol. 61, 125-12. Manickan, E., Yu, Z., Rouse, R.J., Wire, W.S. & Rouse, B.T. (1995) Induction of protective immunity against herpes simplex virus with DNA encoding the immediate early protein ICP 27 Vir. Immunol. 8, 53-61. Michel, M.L., Davis, H.L., Schleef, M., Mancini, M., Tiollais, P. & Whalen, R.G. (1995) DNA-mediated immunization to the hepatitis B surface antigen in mice: aspects of the humoral response mimic hepatitis B viral infection in humans Proc. Natl. Acad. Sci. USA 92, 5307-5311. Moore, M.W., Carbone, F.R. & Bevan, M.J. (1988) Introduction of soluble protein into the class I pathway of antigen processing and presentation Cell 54, 777-785. Mor, G., Klinman, D.M., Shapiro, S., Hagiwara, E., Sedegah, M., Norman, J.A., Hoffman, S.L. & Steinberg, A.D. (1995) Complexity of the cytokine and antibody response elicited by immunizing mice with Plasmodium yoelii circumsporozoite protein plasmid DNA J. Immunol. 155, 2039-2046. Mor, G., Singla, M., Steinberg, A.D., Hoffman, S.L., Okuda, K. & Klinman, D.M. (1997) Do DNA vaccines induce autoimmune disease Hum. Gene Ther. 8, 293-300. Mor, G., Yamshchikov, G., Sedegah, M., Takeno, M., Wang, R., Houghten, R.A., Hoffman, S. & Klinman, D.M. (1996) Induction of neonatal tolerance by plasmid DNA vaccination of mice J Clin. Invest. 98, 2700-5. Nichols, W.W., Ledwith, B.J., Manam, S.V. & Troilo, P.J. (1995) Potential DNA vaccine integration into host cell genome Ann. N. Y. Acad. Sci. 772, 30-39. Pardoll, D.M. & Beckerleg, A.M. (1995) Exposing the immunology of naked DNA vaccines Immunity 3, 165-169. Pertmer, T.M., Roberts, T.R. & Haynes, J.R. (1996) Influenza virus nucleoprotein-specific immunoglobulin G subclass and cytokine responses elicited by DNA vaccination are dependent on the route of vector DNA delivery J. Virol. 70, 6119-6125. Petty, R.E. & Steward, M.W. (1976) The effects of immunological adjuvants on the relative affinity of anti-protein antibodies Immunol. 32, 49-55. Petty, R.E., Steward, M.W. & Soothill, J.F. (1972) The heterogeneity of antibody affinity in inbred mice and its possible immunopathologic significance Clin. Exp. Immunol. 12, 231-241. Prince, A.M., Whalen, R. & B., B. (1997) Successful nucleic acid based immunization of newborn chimpanzees against hepatitis B virus Vaccine 15, 916-919. Raz, E., Carson, D.A., Parker, S.E., Parr, T.B., Abai, A.M., Aichinger, G., Gromkowski, S.H., Singh, M., Lew, D., Yankauckas, M.A. & et al. (1994) Intradermal gene immunization: the possible role of DNA uptake in the induction of cellular immunity to viruses Proc. Natl. Acad. Sci. USA 91, 9519-9523. Raz, E., Tighe, H., Sato, Y., Corr, M., Dudler, J.A., Roman, M., Swain, S.L., Spiegelberg, H.L. & Carson, D.A. (1996) Preferential induction of a Th1 immune response and inhibition of specific IgE antibody formation by plasmid DNA immunization Proc. Natl. Acad. Sci. USA 93, 5141-5145. Roman, M., Martin-Orozco, E., Goodman, J.S., Nguyen, M.D., Sato, Y., Ronaghy, A., Kornbluth, R.S., Richman, D.D., Carson, D.A. & Raz, E. (1997) Immunostimulatory DNA sequences function as T helper-1-promoting adjuvants Nature Med. 3, 849-854. Sato, Y., Roman, M., Tighe, H., Lee, D., Corr, M., Nguyen, M.D., Silverman, G.J., Lotz, M., Carson, D.A. & Raz, E. (1996) Immunostimulatory DNA sequences necessary for effective intradermal gene immunization Science 273, 352-354. Stacey, K.J., Sweet, M.J. & Hume, D.A. (1996) Macrophages ingest and are activated by bacterial DNA J. Immunol. 157, 2116-2122. Ulmer, J.B., Deck, R.R., Dewitt, C.M., Donnelly, J.J. & Liu, M.A. (1996) Generation of MHC class I-restricted cytotoxic T lymphocytes by expression of a viral protein in muscle cells: antigen presentation by non-muscle cells Immunol. 89, 59-67. Ulmer, J.B., Donnelly, J.J., Parker, S.E., Rhodes, G.H., Felgner, P.L., Dwarki, V.J., Gromkowski, S.H., Deck, R.R., De Witt, C.M., Friedman, A. & et al. (1993) Heterologous protection against influenza by injection of DNA encoding a viral protein Science 259, 1745-1749. Wicks, I.P., Howell, M.L., Hancock, T., Kohsaka, H., Olee, T. & Carson, D.A. (1995) Bacterial lipopolysaccharide copurifies with plasmid DNA: implications for animal models and human gene therapy Hum. Gene Ther. 6, 317-323. Wolff, J.A., Ludtke, J.J., Acsadi, G., Williams, P. & Jani, A. (1992) Long-term persistence of plasmid DNA and foreign gene expression in mouse muscle Hum. Mol. Genet. 1, 363-369. Wolff, J.A., Malone, R.W., Williams, P., Chong, W., Acsadi, G., Jani, A. & Felgner, P.L. (1990) Direct gene transfer into mouse muscle in vivo Science 247, 1465-1468. Wolff, J.A., Williams, P., Acsadi, G., Jiao, S., Jani, A. & Chong, W. (1991) Conditions affecting direct gene transfer into rodent muscle in vivo Biotechniques 11, 474-485. Xiang, Z. & Ertl, H.C. (1995) Manipulation of the immune response to a plasmid-encoded viral antigen by coinoculation with plasmids expressing cytokines Immunity 2, 129-135. Yi, A.K., Klinman, D.M., Martin, T.L., Matson, S. & Krieg, A.M. (1996) Rapid immune activation by CpG motifs in bacterial DNA. Systemic induction of IL-6 transcription through an antioxidant-sensitive pathway J. Immunol. 157, 5394-5402. Copyright 1998 Australian Biotechnology Association Ltd. The following images related to this document are available:Line drawing images[au98023a.gif] | ||||||||||||||||||||||||||||||||||||||
| |||||||||
{kind=link}