 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Cloning and Purification of the Hydrophilic
Fragment Gillian Martínez,1 Ariel Viña,1 José L García,2 Juan Morales Grillo1 1Centro de Ingeniería Genética y Biotecnología.
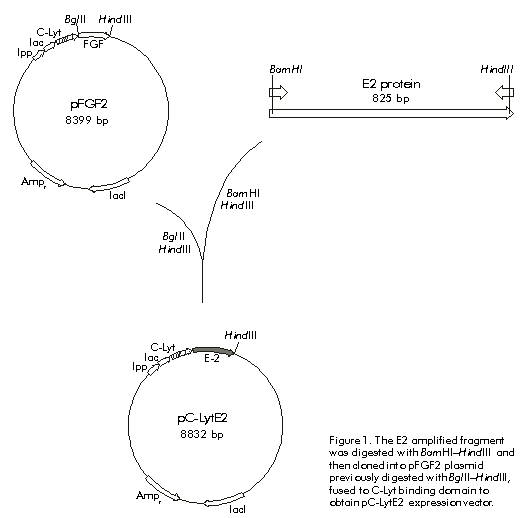
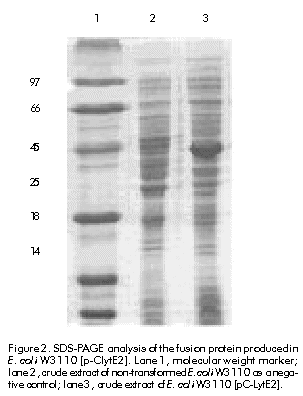
Received in January, 1999. Accepted for publication in August, 1999. Code Number: be00020 Abstract The hydrophilic fragment of hepatitis C virus (HCV) E2 protein was expressed in Escherichia coli as a chimera, bound to the choline-binding domain (C-Lyt) of the major autolysin of Streptococcus pneumoniae. The C-LytE2 chimera was purified by affinity chromatography using DEAE-Sepharose. The purified antigen was used to immunize rabbits and the specific humoral immune response to E2 protein was examined. Induction to high levels of antibodies against this HCV E2 protein was found. Keywords: E2 protein, HCV, humoral immune response, purification Resumen Clonación y purificación del fragmento hidrofílico de la proteína E2 del virus de la hepatitis C, fusionado al dominio de unión a colina de la principal autolisina de Streptococcus pneumoniae (C-Lyt): evaluación de la respuesta inmune humoral en conejos. El fragmento hidrofílico de la proteína E2 del virus de la hepatitis C (VHC), fue expresado en Escherichia coli como una quimera fusionado al dominio C-Lyt. La proteína quimérica C-LytE2 fue purificada por cromatografía de afinidad mediante el uso de DEAE-Sepharosa. Este antígeno purificado se utilizó en la inmunización de conejos y se evaluó la respuesta inmune humoral específica. Se encontró una alta inducción de anticuerpos contra la proteína estructural E2 del VHC. Palabras claves: proteína E2, purificación, respuesta inmune humoral, VHC Introduction Hepatitis C virus (HCV) is the major causative agent of post-transfusion-associated non-A, non-B hepatitis throughout the world [1]. This virus has a 9.4 kb single-stranded positive-sense RNA genome that encodes a polyprotein precursor, which is cleaved post-translationally into putative structural and non-structural proteins [2]. The structural E2 glycoprotein is likely located at the surface of the HCV virion. This association with the surface of the virus makes this protein a major target for humoral immune responses to infection by HCV [3]. Antigenic sequences found within hypervariable region 1 (HVR-1) of the E2 protein have been suggested to act as neutralizing epitopes. However, the emergence of escape mutants makes difficult to exploit this finding for therapeutic purposes [4]. Other antigenic regions outside the HVR-1 have been also identified and characterized [5]. Nevertheless, few data are available regarding the mapping of B cell epitopes in these regions [6. 9]. In vitro neutralization of HCV by antibodies directed to some of these epitopes, has been described [10]. These findings made E2 protein an attractive candidate to study the neutralization of HCV by antibodies directed against this protein, as well as to evaluate its use as a putative vaccine. The aim of this work was to purify the E2 protein in a single step, in order to prepare anti-E2 antibodies that could be useful to perform protein expression experiments in eukaryotic cells. The hydrophilic fragment of E2 protein was expressed in Escherichia coli as a fusion protein containing the choline-binding domain (C-Lyt) of the major autolysin of Streptococcus pneumoniae (C-LytE2). The antigen obtained was purified using DEAE-Sepharose. The purified protein was used for immunization experiments. Materials and Methods Cloning of HCV E2 protein coding sequence A DNA fragment encoding a region of the HCV E2 protein from amino acids (aa) 384 to 650 of the viral polyprotein, was amplified by polymerase chain reaction (PCR). The amplification was performed using the plasmid pHCV as template [11], which contains the cDNA sequence of HCV genome. The oligonucleotides 5'-GGGGATCCTGCAGATACCCACGTGACAGGTGGTGCT-3' (containing a BglII site) and 5'-GGAATTCAAAGCTTATCCTCGAGTCCARTTGCATGCAGC-3' (containing a HindIII site), were used as primers. It is worth noting that a stop codon was introduced by PCR at the 3' end of the coding region. The amplified segment was cloned between the sites HindIII.BglII of pFGF2 plasmid. This plasmid was used as a source of the C-Lyt domain, since it contains the sequence of acidic fibroblast growth factor fused to this domain [12]. The vector also contains a modified E. coli lipoprotein promoter/lactose promoter-operator region (lpp/lac) that can be used to express the C-LytE2 chimeric protein. Figure 1 shows the strategy followed to construct the plasmid pC-LytE2. Expression of the C-LytE2 fusion protein The constructed pC-LytE2 plasmid was transformed into E. coli W3110 strain. A single transformant colony was grown at 37 ºC for 12. 14 h in Luria broth (LB) medium containing ampicillin (50 mg/mL). Cells were diluted in 300 mL of fresh LB medium containing ampicillin and were grown at 37 ºC with vigorous shaking (2500 rpm in thermostat shaking ZT-01, Santiago de Cuba, Cuba), until the culture reached an optical density (OD) of 0.5. 0.8 at 600 nm. The expression of the fusion gene was induced by adding 0.5 mM of isopropyl-b-d-thiogalactopyranoside (IPTG) to the culture medium. After 5 h of induction, the cell extract was analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) according to Laemmli [13]. Western blotting Analysis of proteins bound to polyvinylidene difluoride membranes (PVDF) was performed according to the method of Towbin et al. [14]. The sheet with the transferred proteins was treated for 1 h at room temperature with phosphate-buffered saline (PBS, 0.1 M NaCl, 2 mM KCl, 10 mM Na2HPO4, 1 mM KH2PO4, pH 7.2) containing 5% skim milk. The membrane was then incubated using as the primary antibody either anti-E. coli antibody-free serum [15] from an HCV-infected patient diluted 1:10, or rabbit anti-E2 protein diluted 1:500. After washing with PBS, the membrane was incubated with anti-human immunoglobulin or anti-rabbit immunoglobulin conjugated to horseradish peroxidase (Boehringer-Mannheim, Germany) diluted 1:25,000 as the secondary antibody. After washing, immunoreactivity was detected by enhanced chemoluminescence (ECL, Amersham Pharmacia Biotech, Sweden) according to manufacturer. s instructions. The expression level was determined by densitometry using a molecular analysis program (CS-9000, Shimadzu and Kyoto, Japan). Purification of C-LytE2 fusion protein A modular fermentation system (BE Marubishi, Japan) with a working volume of 1.5 L was used to culture the engineered bacterium. Late-log phase E. coli W3110 [pC-LytE2] cells were harvested by centrifugation at 6000 g for 20 min after induction with IPTG. The resulting pellet was washed once with 10 mM Tris-HCl pH 8.0 containing 6 mM EDTA (TE buffer). The cells were disrupted using a French press (1500 kg cm-2) at 1 g (wet wt) mL-1 in TE. The insoluble material was collected by centrifugation (18 000 xg for 30 min) and washed successively with 1% Triton X-100 in TE and 2 M guanidinium hydrochloride in TE. The fusion protein was solubilized with extraction buffer (0.5 M Tris-HCl pH 8.0, 6 M guanidinium hydrochloride, 0.5 M NaCl, 2 mM DTT) for 2 h by shaking at 4 ºC, and was refolded by diluting ten-fold with renaturing buffer (50 mM Tris-HCl pH 8.0, 50 mM NaCl, 80 mM CuSO4) at 4 ºC for 10 h. Finally, the sample was loaded onto a DEAE-Sepharose column and washed using a gradient of NaCl (0.5. 1.5 M), and then was eluted with a buffer (50 mM Tris-HCl pH 8.0, 1.5 M NaCl) containing 2% choline as described by Ortega et al. [12]. Immunization For primary immunization, two rabbits were injected subcutaneously with an emulsion of equal volumes of PBS containing 100 µg of the purified C-LytE2 protein, and complete Freund. s adjuvant. Two weeks later, the rabbits were boosted by injecting an emulsion of equal volumes of PBS containing the C-LytE2 protein and incomplete Freund. s adjuvant. This treatment was repeated once more. Blood samples were collected after the second booster immunization. Indirect ELISA Microtiter plates (Nunc Immuno Plates, Maxisorp, Costar, Cambridge, USA) were coated for 1 h at 37 ºC with 100 µL of 6 µg/mL antigen (C-LytE2) or 5 mg/mL internal synthetic peptide of the E2 protein spanning aa 545. 570 of the viral HCV polyprotein (NH2-PPQGNWFGCTWMNSTGFTKTCGGPPC-COOH), or 6 µg/mL His-E2 protein in coating buffer (0.05 M carbonate buffer, pH 9.5, 0.02% NaN3). The latter is another variant of E2 protein expressed in E. coli fused to a 6 x His tag spanning aa 458. 680 of the HCV polyprotein (manuscript in preparation). After washing the plates with 0.05% Tween 20 in PBS (PBST), they were incubated in 200 µL of blocking solution containing 5% skim milk in PBST, for 1 h at 37 ºC. Immune serum was diluted in 5% skim milk/PBST, and 100-µL samples were added to the wells. Plates were incubated at 37 ºC for 1 h and then washed five times in PBST. Then, the wells were incubated with anti-rabbit IgG goat Fab conjugated with horseradish peroxidase diluted 1:3000 (Amersham Phramacia Biotech, Sweden), for 1 h at 37 ºC. Plates were finally washed five times in PBST and incubated with a solution containing 100 µL of o-phenylenediamine (OPD) (Sigma, St. Louis, USA) and 0.02% H2O2, for 15 min at room temperature. The reaction was stopped with 50 µL of 2 M H2SO4 and OD was measured at 492 nm in a SensIdent Scan (Merck, USA). Results and Discussion Cloning and expression A DNA fragment containing the E2 protein region (aa 384. 650 of the polyprotein) not including the N- and C-terminal hydrophobic regions, was amplified from pHCV plasmid by PCR. After digesting the amplified fragment with BamHI and HindIII endonucleases, it was cloned into the plasmid pFGF2 previously digested with BglII and HindIII, to obtain the pC-LytE2 expression vector that contains the E2 fragment fused to the C-Lyt domain (Figure 1). Figure 1. The E2 amplified fragment was digested with BamHI–HindIII and then cloned into pFGF2 plasmid previously digested with BglII–HindIII, fused to C-Lyt binding domain to obtain pC-LytE2 expression vector. To produce the quimeric C-LytE2 protein, E. coli W3110 cells transformed with pC-LytE2 were induced with 0.5 mM IPTG as previously described. When the cell extracts were analyzed by SDS-PAGE and the proteins were visualized by staining with Coomassie blue, a prominent band was observed in the induced cultures (Figure 2). The mobility of the induced proteins in denaturing polyacrylamide gels was in agreement with the predicted value (~42 kD), which corresponds to the mass of E2 plus C-Lyt domain. The expression level of C-LytE2 protein was between 8% and 10% of the total protein. A similar expression level has been previously reported for the production of CPL-1 lysozyme using the same expression system [16]. In addition, this E2 fragment has been expressed to similar levels fused to the maltose binding protein in E. coli [17]. Surprisingly, most of the chimeric protein was associated with cellular debris (data not shown). Therefore, considering the high levels of C-LytE2 protein found in association with the cellular debris in this case, it was necessary to extract and refold the insoluble chimeric protein before purification by the C-Lyt system. Figure 2. SDS-PAGE analysis of the fusion protein produced in E. coli W3110 [p-ClytE2]. Lane 1, molecular weight marker; lane 2, crude extract of non-transformed E. coli W3110 as a negative control; lane 3, crude extract of E. coli W3110 [pC-LytE2]. Single-step purification of C-LytE2 fusion protein The single-step purification system based on the C-Lyt domain of the major autolysin of S. pneumoniae works with a high efficiency and specificity [18]. Diethylaminoethanol (DEAE) behaves as a choline analogue; thus, the proteins fused to the C-Lyt domain specifically bind to DEAE-containing matrices [19]. Therefore, this system was used to perform a rapid and simple purification of the C-LytE2 chimeric protein. The C-LytE2 protein was extracted from cellular debris and refolded by slow dilution. The refolded C-LytE2 protein was then purified using a DEAE-Sepharose column (Figure 3). The recovery of this process was approximately 18%. The purity of the C-LytE2 antigen was about 92% as determined by densitometry. The high level of purity has been also obtained for other proteins using this system [18]. Figure 3. A) SDS-PAGE analysis of purified C-LytE2 fusion protein. Lane 1, molecular weight marker; lane 2, protein extract of refolded sample after solubilization; lane 3, one-step purified fusion protein. B) Western blot analysis of purified protein using serum of an HCV infected patient. Lane 1, negative control; lane 2, purified C-LytE2 protein. Western blot analysis using anti-E. coli antibody free serum from an HCV-infected patient, showed the specific recognition of the purified protein (Figure 3B). Minor protein bands, which are smaller in size than the predicted, were also observed. Presumably, these proteins represent a degradation product obtained during the preparation of C-LytE2 protein. Anti-E2 antibody in immunized rabbits The purified C-LytE2 protein was used to immunize rabbits. After a second booster immunization, blood samples were collected. Rabbits showed a high induction of antibodies against this protein (Figure 4A). When the humoral immune response against C-LytE2 antigen was examined by ELISA, a titer of 1:51,200 was found. Figure 4. Recognition by ELISA of the recombinant C-LytE2, the His-E2 proteins (panel A), and the synthetic peptide (P1) of HCV E2 protein (panel B). For determining the antibody titers, serial dilutions of anti-serum were incubated in antigen-coated wells. The antibody titer was calculated as two-fold the average of the preimmune serum value (negative control). Taking into account that the C-LytE2 antigen used for immunization contains the C-Lyt domain, additional ELISA experiments were performed to ensure that the rabbit serum contained specific antibodies against the E2 protein. Plates were coated with an internal E2 synthetic peptide (P1) or another HCV-E2 variant spanning aa 458. 680 that has been partially purified (His-E2) (data not shown). These analyses demonstrated that the rabbit serum recognized the P1 linear epitope with a titer of 1:1000, and the His-E2 protein with a higher titer of 1:25,600 (Figures 4A and 4B). Conclusions A HCV E2 truncated protein spanning aa 384. 680 was cloned bound to the C-Lyt pneumococcal domain and produced in E. coli. The protein was purified by affinity chromatography using DEAE-Sepharose and a high induction of specific antibodies against the E2 protein was found in rabbits immunized with the C-LytE2 fusion protein. References 1. Choo Q-L, Kuo G, Weiner Aj, Overby LR, Bradley DW, Houghton M. Isolation of a cDNA clone derived from a blood-borne non-A, non-B viral hepatitis genome. Science 1989;244:359. 62. 2. Houghton M, Weiner J, Han J, Kuo G, Choo QL. Molecular biology of the hepatitis C virus: implications for diagnosis, development and control of viral disease. Hepatology 1991;14:381. 8. 3. Lesniewski R, Okasinski G, Carric C, Sant V, Desai S, Johnson R, et al. Antibody to hepatitis C virus second envelope (HCV-E2) glycoprotein: a new marker of HCV infection closely associated with viremia. J Med Virol 1995;45:415. 22. 4. Kato N, Sekiya H, Ootsuyama Y, Nakazawa T, Hijikata M, Ohkoshi S, et al. Humoral immune response to hypervariable region 1 of the putative envelope glycoprotein (gp70) of hepatitis C virus. J Virol 1993;67:3923. 30. 5. Zhang ZX, Sönnerborg A, Sällberg M. Antigenic structure of the hepatitis C virus envelope 2 protein. Clin Exp Immunol 1994;98:382. 7. 6. Mink MA, Benichou S, Madaule P, Tiollais P, Prince AM, Inchauspe G. Characterization and mapping of B-cell immunogenic domain in hepatitis C virus glycoprotein using a yeast peptide library. Virology 1994;200:246. 55. 7. Nakano I, Maerfens G, Major ME, Vitvitski L, Dubulsson J, Fournillier A, et al. Immunization with plasmid DNA encoding hepatitis C virus envelope E2 antigenic domains induces antibodies whose immune reactivity is linked to the injection mode. J Virol 1997;71:7101. 9. 8. Tedeschi V, Akatsuka T, Shih JW-K, Bttegay M, Feinstone SM. A specific antibody response to HCV E2 elicited in mice by intramuscular inoculation of plasmid DNA containing coding sequences for E2. Hepatology 1997;25:459. 62. 9. Woo J, Kim K, Jung S, Jeong K, Choi E, Sung Y, et al. Identification of a domain containing B cell epitopes in hepatitis C virus E2 glycoprotein by using mouse monoclonal antibodies. J Virol 1999; 73:11. 5. 10. Shimizu YK, Hijikata M, Iwamoto A, Alter HJ, Purcell RH, Yoshikura H. Neutralizing antibodies against hepatitis C virus and the emergence of neutralization escape mutant viruses. J Virol 1994;68: 1494. 500. 11. Takamizawa A, Mori C, Fuke I, Manabe S, Murakami S, Fujita J, et al. Structure and organization of the Hepatitis C virus genome isolated from human carriers. J Virol 1991;65:1105. 13. 12. Ortega S, García JL, Zaso M, Varela J, Muños-Willery I, Cuevas P, et al. Single-step purification on DEAE-Sephacel of recombinant polypeptides produced in Escherichia coli. Biotechnology 1992;10:795. 8. 13. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970;227: 680. 5. 14. Towbin H, Staehlin T, Gordon J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci USA 1979;76:4350. 4. 15. Maniatis T, Fritsch EF, Sambrook J. Molecular cloning. A laboratory manual. New York: Cold Spring Harbor Laboratory Press; 1982. 16. Sanz JM, García JL. Structural studies of the lysozyme coded by the pneumococcal phage Cp-1. Conformational changes induced by choline. Eur J Biochem 1990; 187:409. 16. 17. Mita E, Hayashi N, Ueda K, Kasahara A, Fusamoto H, Takamizawa A, et al. Expression of MBP-HCV NS1/E2 fusion protein in E. coli and detection of anti-NS1/E2 antibody in type C chronic liver disease. Biochem Biophys Res Comm 1992;183: 925. 30. 18. Sánchez-Puelles JM, Sanz JM, García JL, García E. Immobilization and single-step purification of fusion proteins using DEAE-cellulose. Eur J Biochem 1992;203: 153. 9. 19. Sanz JM, López R, García JL. Structural requirements for conversion of pneumcoccal amidase. A new single-step procedure for purification of this autolysin. FEBS Lett 1988;232:308. 12. Copyright Elfos Scientia 2000 The following images related to this document are available:Line drawing images[ba00020b.gif] [ba00020a.gif] [ba00020c.gif] [ba00020d.gif] |
| |||||||||

{kind=link}
{kind=link}