 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnología Aplicada 2000;17:89-93 Purification of the Recombinant Protein TmpA from Treponema pallidum Using Immobilized Metal Ion Affinity Chromatography, and Its Use in the Serodiagnosis of Syphilis Daniel O Palenzuela,1 Juan M Rivera,1 Boris Acevedo,1 Alina Miranda,1
1División de Inmunotecnología y Diagnóstico.
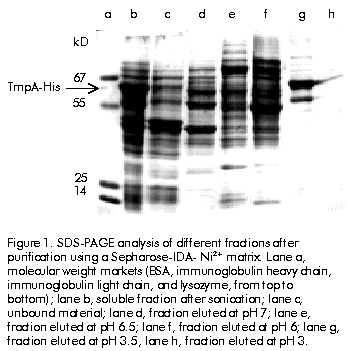
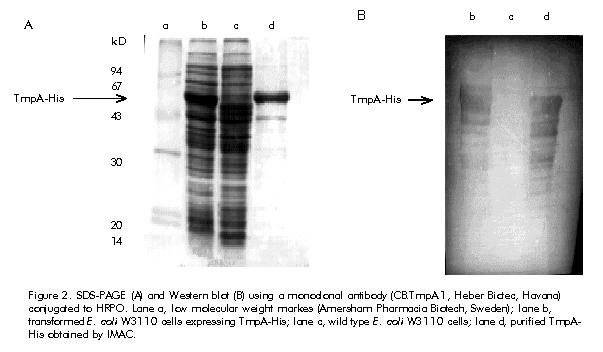
Received in April, 1999. Accepted for publication in October, 1999. Code Number: ba00021 ABSTRACT Keywords: affinity chromatography, diagnosis, IMAC, syphilis, TmpA RESUMEN Palabras claves: cromatografía de afinidad, diagnóstico, IMAC, sífilis, TmpA Introduction Syphilis remains to be a significant cause of morbidity in many developing countries and in some areas of North America and Europe [1]. Treponema pallidum has a moderate to high probability of transmission during contact between susceptible and infected sexual partners. Combined with the fact that treating infected people early in the primary stage of infection reduces the effective period of transmission, this emphasizes the benefits of early accurate serodiagnosis of syphilis [2] Tests used for the serological diagnosis of syphilis can be divided into two categories: nontreponemal and treponemal. Nontreponemal tests (e.g. the venereal disease research laboratory [VDRL] test) are based on a complex antigen mixture composed of an alcoholic solution of cardiolipin, cholesterol, and purified lecitin [3]. Due to reasons still largely unknown, infection with T. pallidum elicits antibodies that react with this antigen mixture. Borkhardt et al. [4] investigated the biological role of antibodies to cardiolipin during T. pallidum infection and suggested that production of antibodies to cardiolipin should be considered a virulence mechanism of pathogenic treponemes with the purpose of evading the host defense mechanisms. The nontreponemal tests have the advantages of being widely available, inexpensive, convenient to perform screening on a large number of specimens, and suitable for determining the efficacy of treatment. However, they are not devoid of limitations, which include the lack of sensitivity in early dark-field-positive primary and late syphilis cases, and the presence of about 1–2% of false-positive individuals among the normal population [5]. Treponemal tests for the diagnosis of syphilis are based on the detection of antibodies directed against T. pallidum antigens elicited after infection. The T. pallidum hemagglutination assay (TPHA) developed in the mid 60’s [6, 7] is widely used in Europe as a screening test, regularly in combination with a nontreponemal assay and also as a confirmatory assay. In some cases, a small percentage of false positive results may be obtained [8]. Recently, a new latex agglutination test (Syphilis Fast, Edinburg University Medical School, UK) and a novel immune-capture enzyme immunoassay (ICE Syphilis Murex Diagnostics, UK) based on the use of three recombinant T. pallidum antigens (TpN15, TpN17 and TpN47), were compared with the native antigen T. pallidum EIA (Captia SelectSyph, Centocor, USA) and VDRL tests. The specificity of Syphilis Fast (99.8%) was significantly higher (p < 0.02) than that of Captia SelectSyph (99.2%) and VDRL (99.1%). There was no difference in the sensitivity of Syphilis Fast and Captia SelectSyph and both were significantly more sensitive than VDRL test. On the other hand, the sensitivity and specificity of ICE Syphilis was significantly higher than that of Captia SelectSyph. These results demonstrated that immunoassays based on recombinant T. pallidum antigens are more specific, as sensitive as the assay based on native antigen, and can be considered a front-line screening test [9, 10]. Among the treponemal antigens that have been expressed by recombinant DNA-techniques, the 42-kD T. pallidum membrane protein A (TmpA) appears to be a promising candidate for mass serodiagnosis of syphilis, as well as a good marker of post-treatment recovery [11, 12]. In the first report regarding the expression of the TmpA protein in Escherichia coli, its purification involved a series of laborious steps that compromised the overall yield [11, 13]. In this paper, we report the purification of a recombinant TmpA bearing a C-terminal affinity tag of six histidine residues (TmpA-His), by a simple one-step method based on immobilized metal ion affinity chromatography (IMAC). Since its first report in 1975 [14], IMAC has become an important tool for the isolation of many proteins and has been widely applied to the purification of recombinant proteins bearing multi-histidine domains at either N- or C-terminus [15, 16]. IMAC has also been applied to purification of single-stranded DNA molecules incorporating a tag consisting of six successive histaminylpurines, which endows a DNA strand with selective retention onto a Ni 2+-NTA-agarose chromatographic matrix [17]. In this work, TmpA-His and a recombinant TmpA variant lacking the six-His tag—the latter purified by electroelution—were compared to VDRL test with respect to their diagnostic performance using an ELISA procedure. The results suggest that TmpA-His protein can be employed in the development of a variety of diagnostic tests for syphilis. Materials and Methods Cloning of TmpA and TmpA-His in E. coli Details about the cloning of the TmpA gene were discussed by Miranda et al. [18]. Briefly, polymerase chain reaction (PCR) was used to isolate the TmpA encoding gene using purified DNA from the lymph of a chancroid lesion of a syphilis patient. The amplified DNA was digested and ligated into the bacterial expression vector pPF-15 [19]. To incorporate a 6-His tag to the TmpA protein at its C-terminus, the TmpA gene was cloned into the pR2M6 vector, a modification of pPF-15 with a 6-His encoding domain 3’ to the inserted gene. Expression of TmpA and TmpA-His in E. coli Both TmpA and TmpA-His were expressed as soluble cytoplasmic proteins in W3110 cells. Cells transformed with the pPF-15 vector containing the TmpA gene were cultured in 5 mL of LB medium [20] supplemented with ampicillin (50 µg/mL) and tryptophan (100 µg/mL), for 12 h at 37 ºC. When cell culture reached an optical density of 1.0 at 610 nm, 5 mL of the culture were inoculated into 500 mL of M9 medium [20] supplemented with 1% casein hydrolysate and 0.2 M glucose, and incubated at 37 ºC for 2 h. Thereafter, b-indole acrylic acid (40 µg/mL) was added and the culture was incubated for 8 h at 37 ºC. Finally, the cells were harvested by centrifugation. E. coli W3110 cells transformed with the pR2M6-TmpA-His vector were grown in M9 medium supplemented with glucose (0.2%), tryptophan (2.4 µg/mL), and ampicillin (50 µg/mL). Cultivation was performed in a 5-L fermenter (B.E. Marubishi, Japan) at 37 ºC for 12 h. Finally, the cells were collected by centrifugation. Purification of TmpA by preparative gel electrophoresis Transformed E. coli cells (1 g) induced for TmpA expression were suspended in 10 mL of 10 mM Tris-HCl pH 8 and 1 mM EDTA. The suspension was sonicated (3 x 150 W burst, 5 ºC) and then clarified by centrifugation (20 min; 2600 xg ; 10 ºC). Two milliliters of supernatant were mixed with 2 mL of sample buffer consisting of 4% SDS, 125 mM Tris-HCl pH 6.9, 20% glycerol, 10% ß-mercaptoethanol, and 0.002% bromophenol blue, and the mixture was boiled for 5 min. This sample was applied onto 3 mm-thick 12.5% SDS-polyacrylamide gel (17 cm high x 17 cm wide). The SDS-PAGE was perfomed as described by Laemmli [21]. The electrophoresis was performed at 10 mA for 12 h. After electrophoresis, the gels were reverse-stained with 0.2 M imidazole and 0.2 M zinc sulfate [22]. The TmpA bands were identified and cut off, and the gel slices introduced into a dialysis bag with 2 mL of electroelution buffer (0.92 M Tris-HCL, 0.2 M glycine, pH 8.3). The dialysis bags were placed into a horizontal electrophoresis chamber and electroeluted at 150 V during 5 h. Preparation of Sepharose 4B-IDA The preparation of iminodiacetic (IDA) Sepharose 4B was carried out according to a reported method [14]: oxirane agarose (125 g wet weight) was suspended in 0.2 M Na 2CO3 (100 mL) pH 11 containing 20 g of iminodiacetic acid. The mixture was kept at room temperature for 24 h and the gel was washed with distilled water. Excess groups were blocked with 1.0 M ethanolamine overnight at room temperature (RT). Ni2+-chelate affinity chromatography One gram of wet biomass was suspended and homogenized in 10 mL of buffer A (0.1 M NaH 2PO4, 0.5 M NaCl, 8 M urea, pH 8). The suspension was sonicated for 2 min, and then clarified by centrifugation. Ten millilitres of clear supernatant were loaded onto a Sepharose-IDA-Ni2+ column (1.6 cm x 20 cm, 10 mL bed volume) equilibrated in buffer A. The column was incubated for 1 h at RT in a bascular shaker. The unbound material was washed out with buffer A until the baseline returned to zero. The adsorbed proteins were eluted in buffer A using a descendent pH-step gradient (pH 7–3). The pH was monitored with a flow-through pH meter (Pharmacia Biotecnology Inc., Sweden). Before each chromatographic performance the column was regenerated by washing, using three bed volumes of 100 mM of EDTA, 1 M NaCl, pH 4.5, followed by washing with five bed volumes of distilled water. A 30-mL volume of a 0.1 M NiCl 2·6H2O aqueous solution pH 4 was then applied, followed by washing with 50 mL of 0.1 M sodium acetate pH 4 to remove the excess of Ni2+. The column was then washed with three bed volumes of distilled water, followed by equilibration in buffer A. The flow rate was maintained at 30 cm/h. Protein determination Total protein concentration in the samples was determined by the method of Bradford [23] using BSA as standard. Western blot analysis The Western blot (WB) was basically perfomed by the method of Towbin [24]. The identification of TmpA in each analysed fraction was performed using an anti-TmpA monoclonal antibody (CB.TmpA.1, Heber Biotec S. A., Cuba) conjugated to horseradish peroxidase (HRPO). The signal was revealed using the insoluble substrate 3,3’-diaminobenzidine (Sigma Chemical Co., USA). Immunoassay for syphilis detection In order to find out the optimal concentration for each protein, 96-well microtiter plates (LabSystems Inc., Finland) were coated with different concentrations of the recombinant proteins TmpA or TmpA-His in 0.5 M sodium carbonate buffer, pH 9.6. Coating was carried out at 37 ºC for 3 h. Serum samples (100 µL/well) were diluted 1:20 in phosphate buffer pH 7.7 containing 0.05% Tween 20 (PBST). The plates were incubated for 30 min at 37 ºC, and washed four times with 200 µL/well of PBST. A protein A/HRPO conjugate (Heber Biotec S.A., Cuba; 100 µL/well) was added and the plates were incubated for 30 min at 37 ºC. Finally, the plates were washed four times as above and incubated for 15 min with 100 µL/well of substrate buffer (citrate phosphate buffer pH 5.5, 0.014% H2O2, 0.25% o-phenylenediamine). The reaction was stopped with 50 µL/well of 2.5 M sulfuric acid, and the optical density read at 492 nm (Sensident, LabSystems Inc., Finland). A panel of 50 VDRL-tested positive and 95 negative sera was used for the evaluation of both proteins in the ELISA described above. All samples were collected in collaboration with the Reference Center for Sexually Transmitted Disease, Marianao’s Blood Blank, Havana, Cuba, and kept at -20 ºC until use. Results and Discussion The T. pallidum TmpA membrane protein was originally cloned and expressed in E. coli K-12 strain using the bacteriophage lambda promoter pL, which is controlled by a thermosensitive repressor [11, 13]. The expressed protein amounted to about 20% of total E. coli proteins and the purification procedure included a complex series of steps involving membrane extraction, acid precipitation, ion-exchange chromatography, and finally, purification by preparative gel electrophoresis followed by electroelution. This process results laborious and expensive with a low overall yield. Initially, TmpA gene was cloned and expressed using the E. coli pPF-15 vector [19]. The pPF-15 vector bears a tryptophan promoter and the T4 terminator, and was designed for the production of in vitro diagnostic recombinant antigens as N-terminal fusion proteins with the first 58 amino acids of human interleukin-2 (IL-2). The antigen was purified by the preparative electroelution procedure (see above). Using this procedure, it was not possible to obtain a highly enriched TmpA, with an estimated yield of 1 mg of pure protein per 200 mg of wet biomass. This is a single-step purification procedure that does not include as many steps as the procedure published by Ijsselmuiden et al. [11] and Van Eijk et al. [13]. Nevertheless, it is expensive and time-consuming with a low overall yield. The purpose of this work was to use the recombinant TmpA protein as an enzyme-linked antigen for the serodiagnosis of syphilis, a considerable amount of protein is needed for large-scale production. To substitute this expensive and low yield purification procedure, we designed a simple purification procedure involving IMAC. For this purpose, TmpA protein gene was amplified by PCR, cloned, and expressed with a C-terminal histidine tag from the pR2M6 vector. As with other proteins previously produced fused to the amino-terminal IL-2 domain [25], the expression level was high, approximately 20% of the total E. coli proteins as detected by 15% SDS-PAGE (Figure 1, lane b ). Figure 1. SDS-PAGE analysis of different fractions after purification using a Sepharose-IDA- Ni2+ matrix. Lane a, molecular weight markets (BSA, immunoglobulin heavy chain, immunoglobulin light chain, and lysozyme, from top to bottom); lane b, soluble fraction after sonication; lane c, unbound material; lane d, fraction eluted at pH 7; lane e, fraction eluted at pH 6.5; lane f, fraction eluted at pH 6; lane g, fraction eluted at pH 3.5, lane h, fraction eluted at pH 3. To purify the TmpA-His by IMAC, the first experiments were performed under nondenaturing conditions (e.g., 0.1 M sodium acetate, 0.5 M NaCl pH 8; or 0.1 M NaH 2PO4, 0.5 M NaCl pH 8) due to the soluble nature of TmpA-His. Under these conditions, TmpA-His was unable to interact with the Sepharose-IDA-Ni2+ matrix, probably because the native conformation of this protein does not favor binding of the histidine tag to the chelating metal. This problem was solved using a high concentration of denaturing chaotropic agent (urea) for protein extraction, and maintaining this concentration during the chromatographic performance, as it has been suggested before [26]. After centrifugation, the crude extract was incubated with the Sepharose-IDA-Ni2+ matrix. Proteins can be eluted by drastically lowering buffer pH and, thus, changing the charge of histidine side chains and reducing their capacity of binding to Ni2+. In fact, by carefully adjusting pH in the washing buffers, it is possible to elute most contaminant proteins before the elution of the histidine-tagged protein. These contaminants can be fixed to the resin even in the absence of surface exposed histidine residues, probably by means of its free alpha-amino groups, which has been shown to be an effective binding site for the purification of synthetic peptides by IMAC [27, 28]. Also, chromatography of human IL-1b and other proteins containing a single surface exposed histidine surrounded by several hydrophobic residues, confirmed that such a sequence could serve as an effective metal-binding domain [29]. In the first experiments, TmpA-His began to elute at a pH value below 6 when the descendent-pH step gradient (pH 7, 6, 5, 4, and 3) was used, with some TmpA-His molecules eluting at pH 4. Accordingly, the column was subsequently washed with buffers at pH 7, 6.5 and 6. For maximal recovery of the TmpA-His bound to resin, elution was performed at a fixed pH value (3.5), albeit the effective final pH value of the eluate is approximately 5 (Figure 1). When the purified protein was studied by Western blot with a HRPO-conjugated anti-TmpA monoclonal antibody, a nuer of degradation products were identified (Figure 2), not only in the final purified protein but also in the initial cell extract. The fact that this degradation is present in the initial cell extract even under denaturing conditions as 8 M urea, which could be used as a neutralizing proteolytic agent [15], suggests that this degradation might occur inside the bacterium. Also, weakly bands migrating above TmpA-His were identified by the specific anti-TmpA monoclonal antibody CB.TmpA.1. These bands are not the result of unspecific binding of CB.TmpA.1 since they are not present in the negative control, so they are more likely to be the result of TmpA polymerization. Figure 2. SDS-PAGE (A) and Western blot (B) using a monoclonal antibody (CB.TmpA.1, Heber Biotec, Havana) conjugated to HRPO. Lane a, low molecular weight markes (Amersham Pharmacia Biotech, Sweden); lane b, transformed E. coli W3110 cells expressing TmpA-His; lane c, wild type E. coli W3110 cells; lane d, purified TmpA-His obtained by IMAC. A purity close to 90% was estimated for TmpA using the data obtained by the Molecular Analyst Software (BioRad, USA) taking into account the degradation products described above. From 1 g of wet biomass about 10 to 15 mg of enriched TmpA-His were obtained, which represents a 75% recovery (Table 1). The stability of TmpA-His conserved at 4 ºC in buffer A pH 5 was studied periodically during 10 months by SDS-PAGE and Western blot, which led to the conclusion that the protein preserves its initial electrophoretic pattern and its antigenic properties. Table 1. Purification and recovery of TmpA-His obtained by IMAC.
When the Sepharose-IDA matrix was charged with Cu 2+, TmpA-His protein started to elute at a pH value (pH 4.5) lower than that when the matrix was charged with Ni2+ (pH 5.5). This result agrees well with the data obtained by Su lkowski et al. [30]. They showed that for different proteins the binding strength for different metals decreases in the following order: Cu2+, Ni2+, Co2+, Zn2+. Unfortunately, the binding strength not only increased for TmpA-His, but also for different E. coli proteins present in the sample. Consequently, some E. coli proteins eluted together with TmpA-His compromising the purity of the final product. Therefore, the Sepharose-IDA-Ni2+ matrix was selected for further works. To demonstrate the usefulness of the TmpA-His protein purified by IMAC in the diagnosis of T. pallidum infection, TmpA-His was compared with the electroeluted TmpA in a direct ELISA as described above. A panel of 50 VDRL positive sera and 95 VDRL negative sera was used in the evaluation. The cut-off value of the assay was taken as the mean optical density of 90 negative sera plus two standard deviations. Comparison between TmpA and TmpA-His by ELISA showed the same sensitivity and specificity (94% and 98.9%, respectively) for the two proteins (Table 2). These results demonstrated that there was no difference between both proteins taking into account their behavior in the ELISA test. Three out of 50 sera positive by VDRL that were negative by ELISA, also resulted negative by TPHA (Fujirebio, Japan), which suggests that these sera are VDRL false positives. This result agrees with the data obtained by other authors [9, 10, 31] who demonstrated the lower specificity of VDRL compared with other assays based on T. pallidum recombinant antigens. The three sera negative by ELISA and TPHA that were positive by VDRL, demonstrated that TmpA-His is a highly specific treponemal antigen, a characteristic that is essential in order to minimize the number of false positives [10] and that could also be used in the development of different diagnostic systems for initial screening of syphilis. Finally, the high expression level of proteins using the pR2M6 vector, and its purification by IMAC, offers a convenient means to rapidly purify large quantities of recombinant proteins that could be used in the diagnosis of different infectious diseases. Table 2. Evaluation by ELISA of the recombinant IMAC-purified TmpA-His and electroeluted TmpA, using a panel of 50 VDRL positive sera and 95 VDRL negative sera.
References 1. Podwinska J. Syphilis and AIDS. Arch Immnol Ther Exp Warsz 1996;44:329–33. 2. Garnett GP, Aral SO, Hoyle DV, Cates W Jr, Anderson RM. The natural history of syphilis. Implications for the transmission dynamics and control of infection. Sex Transm Dis 1997;24:185–200. 3. Rudolph AH, Larsen SA. I n: Demis DJ, editor. Laboratory diagnosis of syphilis, Clinical Dermatology. JB Lippincott Co., Philadelphia; 1993. p.16–22. 4. Borkhardt HL, Zielinski S. Influence of cardiolipin antibodies on the binding of treponemal specific antibodies in the fluorescence treponemal antibody absorption test and the T. pallidum immobilisation test. J Med Microbiol 1997;46:965–72. 5. Sandra AL, Steiner BM, Rudolph AH. Laboratory diagnosis and interpretation of test for syphilis. Clinical Microb Rev 1995;8:1. 6. Rathlev T. Haemaglutination test utilising antigens from pathogenic T. pallidum for serodiagnosis of syphilis. Br J Vener Dis 1967;43:181–5. 7. Meyer JCh. Laboratory diagnosis of syphilis. Curr Probl Dermatol 1996; 24:1–11. 8. Wong SS, Teo DL, Chan RK. Confirmatory serological testing of blood donors positive on TPHA screening in Singapore. Int J STD AIDS 1997;8:760–3. 9. Young H, Moyes A, De Ste Croix I, McMillan A. A new recombinant antigen latex agglutination test (Syphilis Fast) for the rapid serological diagnosis of syphilis. Int J STD AIDS 1998;9:196–200. 10. Young H, Moyes A, Seagar L, McMillan A. Novel recombinant antigen enzyme immunoassay for serological diagnosis of syphilis. J Clin Microbiol 1998;36:913–7. 11. Ijsselmuiden OE, Schouls LM, Stolz E, Aelbers GNM, Agterberg CM, Top J, et al. Sensitivity and specificity of an enzyme-linked immnosorbent assay using the recombinant DNA-derived T. pallidum protein TmpA for assessing the effect of antibiotic therapy. J Clin Microbiol 1989; 27:152–7. 12. Stienstra S, Peeters T, Van der Straaten AM, Kadir N. Treponema pallidum membrane protein A ELISA: a new test for screening and diagnosis of syphilis. Transfusionsmedizin 1992;30:85–91. 13. Van Eijk RVW, Van Embden JDA. Molecular characterization of T. pallidum responsible for human immune response to syphilis. J Microbiol 1982;48:486–7. 14. Porath J, Carlsson J, Olsson J, Belfrage G. Metal chelated affinity chromatography, a new approach to protein fractionation. Nature 1975;258:598. 15. Janknech R, De Martynoff G, Lou J, Hipskind RA, Nordheim A, Stunnenberg HGS. Rapid and efficient purification of native histidine-tagged protein expressed by recombinant vaccinia virus. Proc Natl Acad Sci USA 1991;88:8972. 16. Hochuli E, Bannwarth W, Döbeli H, Gentz R, Stüber D. New metal chelated absorbent selective for proteins and peptide containing neighbouring histidine residues. Bio/Technology 1988;6:1321. 17. Min C, Verdine GL. Immobilized metal affinity chromatography of DNA. Nucleic Acids Res 1996;24:3806–10. 18. Miranda A, Benítez J, Acevedo B, Domínguez MC, Villareal A, Palenzuela D, et al. A comparison of VDRL and immunoassays developed with a recombinant TmpA antigen in the screening of antibodies to T. pallidum. Serodiagnosis and Immunotherapy in Infectious Diseases 1997;8:149–155. 19. Novoa LI, Machado J, Fernández JR, Benítez J, Narciandi RE, Reinoso JL, et al. Method for the expression of heterologous proteins procedure in fused form in E. coli, use therefore, expression vectors and recombinant strains. European Patent Application. Application number: 90202108.8. Publication number: 0416673A1. 1990. 20. Maniatis T, Fritsch EF, Sambrook J. Molecular cloning. A laboratory manual, NY: Cold Spring Harbor Laboratory Press;1982. 21. Laemli UK. Cleavage of structural proteins during assembly of the head of bacteriophage T4. Nature 1970;227:680. 22. Fernández PC, Pedro R, Castellanos L. Reverse staining of sodium dodecyl sulfate-polyacrylamide gels by imidazol-zinc salts: sensitive detection of unmodified proteins. Biotechniques 1992;12:564–72. 23. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilising the principle of protein-dye binding. Anal Biochem 1976;72:248. 24. Towbin H, Staehelin T, Gordon J. Electrophoresis transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci USA 1979;76:4350–3. 25. Novoa LI, Machado J, Benítez J, García J, Narciandi RE, Padrón G, et al. Development of a second generation assay for the detection of antibodies against HIV-1. Biotecnología Aplicada 1992;9:140–7. 26. Hutchens TW, Tai-Tung Yip. Protein interaction with surface-immobilized metal ions: structure-dependent variations in affinity and binding capacity with temperature and urea concentration. J Inorganic Biochemistry 1991;42:105. 27. Johnson RD, Todd RJ, Arnold FH. Multipoint binding in metal affinity chromatography II. Effect of pH and imidazole on chromatographic retention of engineered histidine-containing cytochromes c. J Chromatogr A 1996;725:225–35. 28. Hansen P, Anderson L, Lindeberg G. Purification of cysteine-containing synthetic peptides via selective binding of the alpha-amino group to immobilised Cu2+ and Ni2+ ions. J Chromatogr A 1996;723:51–9. 29. Patwardhan AV, Goud GN, Koepsel RR, Ataai MM. Selection of optimum affinity tags from a phage displayed peptide library. Application to immobilized copper (II) affinity chromatography. J Chromatogr A 1997;787:91–100. 30. Sulkowski E, Vastola K, Oleszek D, Von Muenchhausen W. Surface topography of interferons: a probe by metal chelate chromatography. Anal Chem Symp Ser 1982; 9:313. 31. Thakar YS, Chande C, Mahalley AD, Saoji AM. Seroprevalence of syphilis by TPHA test. Indian J Pathol Microbiol 1996;39:135–8. Copyright Elfos Scientiae 2000 The following images related to this document are available:Line drawing images[ba00021a.gif] [ba00021b.gif] | ||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}