 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnología Aplicada 2000;17:94-98 Clonación y caracterización de los genes que
codifican María Araque, Beatriz Nieves, Elsa Velazco, Zoila Caldera Departamento de Microbiología y Parasitología. Facultad de Farmacia.
Universidad de Los Andes. 5101-A, Venezuela. Recibido en septiembre de 1999. Aprobado en enero del 2000. Code Number: ba00022 RESUMEN Se estudiaron 12 cepas de Klebsiella pneumoniae con un patrón inusual de resistencia a las cefalosporinas de amplio espectro y al aztreonam. Estas cepas se aislaron de pacientes neonatos con septicemia nosocomial recluidos en el Hospital Universitario de Los Andes, Mérida, Venezuela. Además de la resistencia a los b-lactámicos, los aislados clínicos también fueron resistentes a gentamicina, kanamicina, tetraciclina y cloramfenicol, pero permanecieron susceptibles a las cefamicinas y al imipenem. Mediante conjugación, se pudo transferir todos los marcadores de resistencia a células de Escherichia coli MKD135 y, posterior al tratamiento con naranja de acridina, se observó que se perdían estos marcadores. El análisis de los transconjugantes por isoelectroenfoque, permitió detectar la actividad de dos b-lactamasas: una de punto isoeléctrico (pI) 5,4 y otra de pI 8,2. Los genes que codifican ambas enzimas fueron localizados en un plásmido autotransferible de 87 kpb (pNU147). Los ensayos de hibridación, clonación y secuenciación demostraron que el gen blaTEM-1, que codifica la b-lactamasa TEM-1, se encuentra en un fragmento EcoRI de 6 kpb proveniente del plásmido pNU147. En otro fragmento EcoRI de 3 kpb del mismo plásmido, se localizó el determinante que media la producción de una b-lactamasa de espectro extenso (bLEE) de tipo SHV-5. Este estudio confirma que el determinante de la bLEE de tipo SHV-5, el de la b-lactamasa TEM-1 y los genes que confieren resistencia a otros antibióticos no relacionados, son transportados por un plásmido conjugativo (pNU147) capaz de diseminarse entre cepas de K. pneumoniae de origen nosocomial. Palabras claves: b- lactamasa de espectro extenso, b-lactamasa SHV-5, b-lactamasa TEM-1, Klebsiella pneumoniae, septicemia nosocomial ABSTRACT Cloning and Characterization of Genes Encoding TEM-1 and SHV-5 b-Lactamases Isolated in Klebsiella pneumoniae Strains >From Neonates With Nosocomial Septicemia. Twelve Klebsiella pneumoniae strains with an unusual pattern of resistance to broad-spectrum cephalosporins and to aztreonam were studied, isolated from neonates with nosocomial septicemia at the in patient facilities of Los Andes University Hospital, Mérida, Venezuela. These strains were also resistant to gentamicin, kanamycin, tetracycline and chloramphenicol, but remained susceptible to cefoxitin and imipenem. All resistance determinants were transferable to Escherichia coli MKD135 by conjugation and were lost en bloc after treatment with acridine orange. All transconjugants showed the activity of two b-lactamases: one at an isoelectric point (pI) of 5.4 and another at a pI close to 8.2. Genes encoding both enzymes are located on the single, self-transferable 87-kpb plasmid pNU147. Hybridization, cloning and sequencing revealed that one 6-kpb EcoRI fragment of pNU147 carries the gene blaTEM-1 encoding the TEM-1 b-lactamase. Another 3-kpb EcoRI fragment of this plasmid showed a blaSHV-type gene encoding the SHV-5 b-lactamase. This study confirms that extended-spectrum-b-lactamase (ESbL) determinants of the SHV-type —TEM-1 b-lactamase— and the genes enconding resistance to other unrelated antibiotics are carried by a conjugative plasmid (pNU147) capable of spreading itself among nosocomial isolates of K. pneumoniae. Keywords: extended-spectrum b-lactamase, Klebsiella pneumoniae,
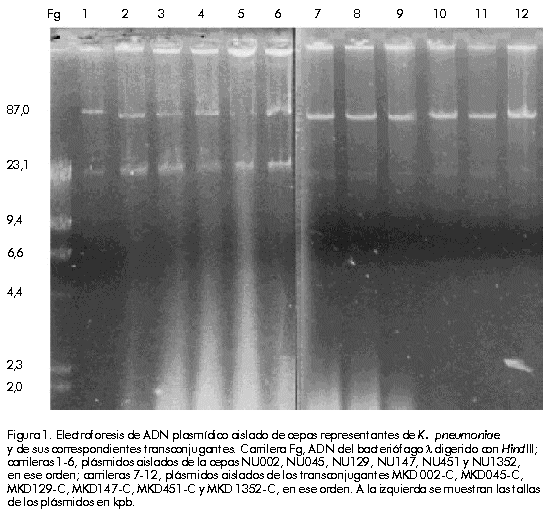
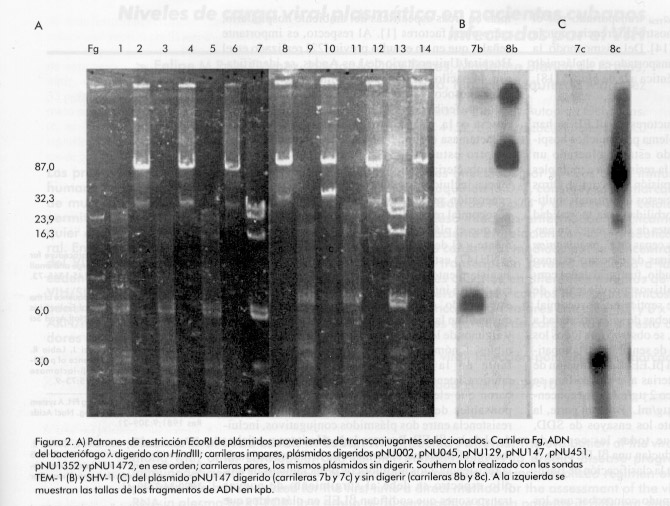
nosocomial septicemia, Introducción En las últimas décadas, la resistencia bacteriana a los antibióticos b-lactámicos ha aumentado notablemente [1]. La causa de este incremento ha sido la aparición y diseminación de b-lactamasas de espectro extenso (bLEE) que hidrolizan, además de los clásicos b-lactámicos, las cefalosporinas de tercera generación y los monobactámicos [2]. Estas enzimas son el resultado de mutaciones en los genes estructurales de las b-lactamasas clásicas TEM-1, TEM-2 y SHV-1 [1, 2]. En 1983, Knothe y colaboradores [3] describieron, por primera vez, la transferencia de los determinantes de resistencia a las cefalosporinas de amplio espectro entre cepas de Klebsiella pneumoniae y Serratia marcescens. Este nuevo fenotipo de resistencia se debe a la producción de una bLEE de tipo SHV-2. Posteriormente, en 1985, se describió en Francia el primer brote epidémico de cepas de K. pneumoniae productoras de bLEE [4]. Desde entonces, se han reportado numerosos brotes causados por organismos productores de bLEE en muchas partes del mundo [1]. Actualmente, entre 10% y 30% de las cepas de K. pneumoniae aisladas en Europa son productoras de bLEE de tipo TEM o del grupo SHV [2]. En este sentido, es de vital importancia la detección de la producción de bLEE en aislados clínicos, no sólo para indicar la antibióticoterapia adecuada a los pacientes infectados con este tipo de cepas, sino también para mantener estos determinantes de resistencia bajo vigilancia epidemiológica [5]. En este estudio, se describe la caracterización genética de un plásmido conjugativo que media la producción de dos b-lactamasas, TEM-1 y SHV-5, y que fue aislado durante un brote epidémico de K. pneumoniae resistente a ceftazidima en una unidad de terapia intensiva neonatal. Materiales y Métodos Cepas bacterianas Durante un brote epidémico que se produjo entre noviembre de 1997 y marzo de 1998 en la Unidad de Terapia Intensiva Neonatal del Hospital Universitario de Los Andes, Mérida, Venezuela, se aislaron 12 cepas de K. pneumoniae con un patrón inusual de resistencia a las cefalosporinas de tercera generación y al aztreonam (monobactámico). Estas cepas fueron recobradas de muestras de sangre de neonatos con septicemia nosocomial e identificadas a través de métodos estandarizados [6]. En la Tabla 1, se resumen las características de las bacterias estudiadas, así como de otras cepas bacterianas y otros plásmidos utilizados en este estudio. Tabla 1. Cepas bacterianas, plásmidos y bacteriófagos utilizados en este estudio.
b LEE, b -lactamasa de espectro extenso; pI, punto isoeléctrico; IEF, isoelectroenfoque;
RIFr, resistencia a Susceptibilidad a los agentes antimicrobianos La concentración inhibitoria mínima (CIM) se determinó por el método de dilución en agar, de acuerdo con las recomendaciones del Comité Nacional de Procedimientos de Laboratorios Clínicos (del inglés, National Committee for Clinical Laboratory Standards, PAIS) [8]. Los antibióticos empleados fueron: cefotaxima (Hoechst Roussel, Somerville, E.U.A.), ceftazidima (Glaxo Group Research Ltd., Greenford, Reino Unido), ácido clavulánico (Beechman, Bristol, E.U.A.), amoxicilina, cefoxitin e imipenem (Merck, Rahway, E.U.A.), aztreonam (Bristol-Myers Squibb, Princenton, E.U.A.), ampicilina, cefalotina, rifampicina, cloramfenicol, tetraciclina, gentamicina y kanamicina (Sigma, St. Louis, E.U.A). La detección inicial de bLEE se realizó mediante la prueba de sinergismo del doble disco (SDD) descrita por Jarlier y colaboradores [9]. Ensayos de conjugación y "curado" de plásmidos Los ensayos de conjugación se llevaron a cabo en medio sólido de acuerdo con la técnica descrita por Willets [10]. Se utilizaron cultivos en fase exponencial mezclados con la cepa receptora E. coli MKD135 en una proporción 1:10. Los transconjugantes fueron seleccionados en medio de Mueller Hinton (Difco, E.U.A.) suplido con ceftazidima 50 mg/mL y rifampicina 400 mg/mL. Los marcadores de resistencia se eliminaron con naranja de acridina según el método descrito por Miller [11]. Detección de b-lactamasas por isoelectroenfoque Estos ensayos se realizaron en geles de poliacrilamida que contenían anfolitos con rango de pH 3-10 (Pharmacia), de acuerdo con el método de Matthew y colaboradores [12]. El isoelectroenfoque (IEF) se realizó en un equipo LKB 2117 MultiphorII (Pharmacia, Suecia)en las siguientes condiciones: 30 W; 1 500 V; 30 mA; durante 4 h a 4 ºC. Para detectar la actividad enzimática, se colocó sobre el gel un papel de filtro saturado con una solución de nitrocefina (Glaxo Group Research Ltd., Greenford, Reino Unido) a 0,25 mM. Técnicas de ADN recombinante Los procedimientos básicos de ADN recombinante se realizaron de acuerdo con Sambrook y colaboradores [13]. Los plásmidos se aislaron por el método de lisis alcalina [14], se separaron por electroforesis en geles de agarosa 0,6% y se tiñeron con bromuro de etidio 0,5 mg/mL. Todos los geles se observaron en una cámara con transiluminación UV y se fotografiaron con película Polaroid tipo 667. El peso molecular de los plásmidos aislados se estimó mediante el programa Gel Analysis Software (Scanalytics Inc., Lincoln, E.U.A.). Las hibridaciones de Southern blot se realizaron con el empleo de sondas de ADN marcadas con 32P mediante la técnica de cebadores al azar o random priming. Las sondas específicas de blaTEM y blaSHV se amplificaron inicialmente a través de la reacción en cadena de la polimerasa (RCP), con los cebadores conocidos para blaTEM-1 [15] y blaSHV-1 [16], respectivamente. La endonucleasa EcoRI se utilizó de acuerdo con las recomendaciones del proveedor (Boehringer Mannheim, Alemania). Secuenciación del ADN Las secuencias de ADN se determinaron para ambas cadenas según el método de terminación descrito por Messing y colaboradores [17] y el protocolo del estuche de reactivos Thermosequenase Cycle Kit (Amersham Pharmacia Biotech, Suecia). Estos ensayos se realizaron en un secuenciador automático de ADN (Modelo 400, LI-COR Inc.). Los cebadores se marcaron con IRD-800 (MWG-Biotech, München, Alemania). Resultados Análisis fenotípico de cepas de K. pneumoniae de origen nosocomial Las 12 cepas de K. pneumoniae mostraron perfiles de susceptibilidad compatibles con la producción de bLEE (datos no mostrados). Los rangos inhibitorios de la ceftazidima sobre las cepas estudiadas, fueron de 2 a 4 veces mayores que la CIM del cefotaxime (32 a 128 mg/mL). Sin embargo, en todos los casos se restituyó la susceptibilidad a la ceftazidima y se alcanzaron rangos inhibitorios < 0,5 mg/mL cuando se empleó el ácido clavulánico a una concentración de 2 mg/mL. La prueba de SDD fue consistente con los resultados obtenidos con la CIM. Además de la resistencia a las cefalosporinas de amplio espectro, se pudo detectar que las cepas de K. pneumoniae presentaban resistencia asociada a la kanamicina, la gentamicina, la tetraciclina y el cloramfenicol. Sin embargo, permanecieron susceptibles al cefoxitin y al imipenem, y mostraron concentraciones inhibitorias < 2 mg/mL. Transferencia conjugativa de los marcadores de resistencia Los genes que confirieron la resistencia a los antibióticos b-lactámicos fueron transferidos por conjugación a células receptoras de E. coli MKD135, con una frecuencia aproximada de 104 transconjugantes por célula donante. Los transconjugantes mostraron un patrón de resistencia semejante al que obtuvieron inicialmente las células donantes. Los determinantes de resistencia a los aminoglucósidos, las tetraciclinas y el cloramfenicol, siempre fueron cotransferidos con la bLEE. La prueba de SDD fue positiva en todos los transconjugantes. El análisis de los extractos crudos de los transconjugantes por IEF, reveló el mismo perfil de actividad enzimática en todos los casos al evidenciarse dos bandas, una con pI 5,4 y otra con pI 8,2 (Tabla 2). Estos resultados sugieren que se transfirieron por conjugación dos determinantes diferentes de b-lactamasas. Análisis del perfil plasmídico Los plásmidos extraídos de las cepas de K. pneumoniae —y de sus correspondientes transconjugantes—, mostraron el mismo perfil plasmídico (Figura 1). De la misma forma, se pudo observar que los patrones de restricción obtenidos fueron indistinguibles entre los plásmidos aislados luego de la digestión con EcoRI (Figura 2A). El peso molecular de los plásmidos se estimó en 87 kpb. El curado de plásmidos con naranja de acridina permitió eliminar todos los marcadores de resistencia en conjunto, lo que indicó que la resistencia a las cefalosporinas de amplio espectro, así como al aztreonam, la gentamicina, la kanamicina, la tetraciclina y el cloramfenicol, es transportada por un mismo plásmido conjugativo. Hibridación de Southern blot La utilización de sondas específicas para la detección de b-lactamasas (TEM-1 y SHV-1) en plásmidos digeridos con EcoRI y sin digerir, permitió evidenciar que un fragmento EcoRI de 3 kpb presentaba secuencias relacionadas con la sonda blaSHV-1, mientras que la señal de hibridación obtenida con la sonda TEM-1 observó indistinguiblemente sobre dos fragmentos EcoRI con longitudes de 6 kpb aproximadamente. En consecuencia, estos dos fragmentos fueron aislados y purificados de forma separada y, posteriormente, sometidos a una segunda hibridación con la sonda TEM-1. Los resultados obtenidos permitieron demostrar que la banda superior más cercana a 6 kpb y la sonda blaTEM-1, tenían secuencias homólogas (Figura2 B, C). Caracterización genética El plásmido pNU147 de la cepa NU147 fue seleccionado como representante del grupo para la caracterización del determinante bLEE. Con este propósito, se clonaron en el vector pBC-SK los fragmentos EcoRI del pNU147 que dieron una señal positiva en las pruebas de hibridación para las dos b-lactamasas diferentes. Estos ensayos produjeron los siguientes plásmidos recombinantes: pBC-147/6, que contiene el fragmento EcoRI de 6 kpb portador del alelo blaTEM, y pBC-147/3, que posee el fragmento EcoRI de 3 kpb portador del alelo blaSHV. Posteriormente, a los clones recombinantes se les determinó el fenotipo de resistencia a los antibióticos b-lactámicos (Tabla 3). E. coli DH5a [pBC-147/6] fue resistente a la ampicilina, la amoxicilina y el cefalotin, y continuó siendo sensible a las cefalosporinas de segunda y tercera generaciones, así como al imipenem y al aztreonam. El análisis de estos clones recombinantes por IEF, permitió detectar la presencia de una b-lactamasa de pI 5,4. Por otro lado, el clon E. coli DH5a [pBC-147/3] mostró un perfil fenotípico compatible con la producción de bLEE de pI 8,2. La secuenciación del alelo blaTEM transportado por el plásmido pBC-147/6, no mostró diferencias con la secuencia del alelo blaTEM-1 [14]. Del mismo modo, la secuencia del alelo blaSHV transportado en el plásmido pBC-147/3 demostró ser idéntica a la de blaSHV-5 [18]. Discusión Los microorganismos productores de bLEE se han convertido en un grave problema para muchos hospitales, especialmente cuando está involucrado un plásmido común que media la resistencia a múltiples antibióticos [1, 5] en la transmisión nosocomial. Otros reportes han confirmado que estos organismos multirresistentes aumentan la morbilidad y la mortalidad significativamente en pacientes de alto riesgo, en particular los neonatos [19]. Las cepas de K. pneumoniae resistentes a las cefalosporinas de espectro extenso que se analizaron en este estudio, fueron aisladas consecutivamente de hemocultivos provenientes de neonatos con diagnóstico de septicemia nosocomial. Cuando se realizaron las pruebas de susceptibilidad a los agentes antimicrobianos, se observó que todos los aislados mostraban un perfil de sensibilidad compatible con la producción de una bLEE. La restitución de la sensibilidad de estas bacterias a la ceftazidima se observó con ácido clavulánico 2 mg/mL, para concentraciones inhibitorias < 0,5 mg/mL. Por otra parte, la detección de bLEE mediante los ensayos de SDD, permitieron confirmar que todas las cepas de K. pneumoniae aisladas producían una bLEE clase A perteneciente al grupo 2be de la clasificación de Bush-Jacoby-Medeiros [20]. En todos los aislados se pudo comprobar que los determinantes para la bLEE, así como los de resistencia a la gentamicina, la kanamicina, la tetraciclina y el cloramfenicol, estaban codificados en un plásmido autotransferible de 87 kpb (pNU147). El fenómeno de la multirresistencia en cepas productoras de bLEE, parece ser una característica común en especies pertenecientes a la familia Enterobacteriaceae [1, 2]. En este sentido, se ha confirmado que las bLEE son codificadas por genes que se encuentran localizados en plásmidos grandes y, por lo general, estos plásmidos son asiento para la inserción de otros genes de resistencia [2, 5]. La caracterización genética y molecular del pNU147 mediante los estudios de hibridación, clonación y secuenciación, reveló que un fragmento EcoRI de 3 kpb transporta el gen blaSHV que codifica una bLEE de tipo SHV-5. De forma interesante, se encontró un gen que codifica otra b-lactamasa, la TEM-1, en un fragmento EcoRI de 6 kpb del mismo. Aunque se han reportado aislados clínicos de Enterobacteriaceae productores de más de un tipo de b-lactamasa del grupo TEM [21], no se ha descrito la presencia simultánea de genes que codifiquen una b-lactamasa de tipo TEM y otra del grupo SHV en un mismo plásmido autotransferible. La evolución de los genes que median la resistencia a las cefalosporinas de amplio espectro y al aztreonam en miembros de la familia Enterobacteriaceae, está directamente relacionada con el uso de los antibióticos y con la aparición de brotes o epidemias en áreas específicas del ambiente hospitalario, entre otros factores [1]. Al respecto, es importante señalar que en un estudio previo [22] realizado en el Hospital Universitario de Los Andes, se identificaron 44 bacilos gramnegativos multirresistentes de origen nosocomial, de los cuales se aisló un plásmido autotransferible de 54 kpb responsable de la resistencia a la gentamicina, y que codifica una b-lactamasa de tipo TEM-1. Poco tiempo después, en otro estudio [23] se identificaron diversas enterobacterias con múltiples marcadores de resistencia, incluidos los de las cefalosporinas de tercera generación, provenientes de pacientes con infección nosocomial recluidos en el mismo hospital. Es posible que el plásmido de 54 kpb identificado previamente y el de 87 kpb caracterizado en este estudio (pNU147) estén relacionados genéticamente, y que su aislamiento en eventos diferentes sea consecuencia del uso intenso de determinados antibióticos. En este sentido, se debe tener en cuenta que los genes que median la producción de la b-lactamasa TEM-1 y algunos de los que codifican las bLEE son transponibles, fenómeno que desempeña un papel importante en la evolución de los plásmidos de multirresistencia [24]. Sirot y colaboradores comprobaron que elementos transponibles fueron los responsables de transferir varios determinantes de resistencia entre dos plásmidos conjugativos, incluido el de las bLEE. Por otro lado, Heritage y colaboradores afirman que el aumento de la diseminación de la resistencia a los antibióticos b-lactámicos de amplio espectro se debe al surgimiento de nuevos transposones que codifican bLEE en plásmidos que circulan en áreas hospitalarias donde el uso de estos antibióticos es intenso. En conclusión, este estudio confirma que circula un plásmido epidémico en la Unidad de Terapia Intensiva Neonatal del Hospital Universitario de Los Andes, Mérida, Venezuela. La utilización de técnicas de ADN recombinante permitió identificar que este plásmido conjugativo de 87 kpb, pNU147, transporta los determinantes que median la producción de una bLEE de tipo SHV-5 y una b-lactamasa de tipo TEM-1, además de otros genes que confieren resistencia a la gentamicina, la kanamicina, la tetraciclina y el cloramfenicol. Por consiguiente, es necesario mantener estas cepas bajo vigilancia epidemiológica y establecer nuevas políticas que aseguren el uso adecuado de los antibióticos en esta institución. No obstante, el conocimiento de los factores moleculares y genéticos involucrados en la generación de resistencia, así como de los elementos que favorecen la transmisión genética de esta información, son puntos claves para implementar estrategias futuras que ayuden a combatir los microorganismos emergentes con resistencia a los agentes antimicrobianos. Agradecimientos Este trabajo fue financiado por el Consejo de Desarrollo Científico, Humanístico y Tecnológico de la Universidad de Los Andes (CDCHT), a través de los proyectos FA192-96-07-AA y FA193-96-07-C. Rereferences 1. Medeiros AA. Evolution and dissemination of b-lactamases accelerated by generations of b-lactam antibiotics. Clin Infect Dis 1997;24 (Suppl 1):19–45. 2. Livermore DM. b-lactamases in laboratory and clinical resistance. Clin Microbiol Rev 1995;8:557–84. 3. Knothe H, Shah P, Krcmery V, Antal M, Mitsuhashi S. Transferable resistance to cefotaxime, cefoxitin, cefamandole and cefuroxime in clinical isolates of Klebsiella pneumoniae and Serratia marcescens. Infection 1983;11:315–7. 4. Sirot D, Sirot J, Labia R, Morand A, Courvalin P, Darfeuille-Michaud A, et al. Transferable resistance to third-generation cephalosporins in clinical isolates of Klebsiella pneumoniae: identification of CTX-1, a novel b-lactamase. J Antimicrob Chemother 1987;20:223–34. 5. Livermore DM. b-Lactamase-mediated resistance and opportunities for its control. J Antimicrob Chemother 1998; 41 (Suppl D):25–41. 6. Farmer III JJ. Enterobacteriaceae: introduction and identification. In: Murray PR, Baron EJ, Pfaller MA, Tenover FC, Yolken RH, editors. Manual of clinical microbiology, Washington: ASM PRESS; 1995, p 438–49. 7. Kholodii GYa, Mindlin SZ, Bass IA, Yurieva, OV, Minakhina SV, Nikiforov VG. Four genes, two ends, and a res region are involved in transposition of Tn5053: a paradigm for a novel family of transposons carrying either a mer operon or an integron. Mol Microbiol 1995;17: 1189–200. 8. National Committee for Clinical Laboratory Standards. Methods for dilution antimicrobial susceptibility test for bacteria that grow aerobically. Approved standard. Villanova, PA, National Committee for Clinical Laboratory Standards, 1997; NCCLS Document M7-A4. 9. Jarlier V, Nicolas M, Fournier G, Philippon A. Extended broad-spectrum b-lactamases conferring transferable resistance to newer b-lactam agents in Enterobacteriaceae: hospital prevalence and susceptibility patterns. Rev Infect Dis 1988;10:867–78. 10. Willets N. Conjugation. Methods Microbiol 1984;17:33–59. 11. Miller J. Experiments in molecular genetic. New York: Cold Spring Harbor Laboratory Press; 1972. 12. Matthew M, Harris AM, Marshall MJ, Ross GW. The use of isoelectric focusing for detection and identification of beta-lactamases. J Gen Microbiol 1975; 88:169–78. 13. Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual. 2nd ed. New York: Cold Spring Harbor Laboratory Press; 1989. 14. Kado CI, Liu ST. Rapid procedure for detection and isolation of large and small plasmids. J Bacteriol 1981; 145:1365–73. 15. Sutcliffe JG. Nucleotide sequence of the ampicillin resistance gene of Escherichia coli plasmid pBR322. Proc Natl Acad Sci USA 1978;75:3737–41. 16. Barthélémy M, Péduzzi J, Labia R. Complete amino acid sequence of p453-plasmid-mediated PIT-2 b-lactamase (SHV-1). Biochem J 1988;25:73–9. 17. Messing J, Crea R, Seeburg PH. A system for shotgun DNA sequencing. Nucl Acids Res 1981;9:309–21. 18. Billot-Klein D, Gutmann L, Collatz E. Nucleotide sequence of the SHV-5 b-lactamase gene of a Klebsiella pneumoniae plasmid. Antimicrob Agents Chemother 1990;34:2439–41. 19. Hart CA. Klebsiellae and neonates. J Hosp Infect 1993;23:83–96. 20. Bush K, Jacoby GA, Medeiros A. A functional classification scheme for b-lactamases and its correlation with molecular structure. Antimicrob Agents Chemother 1995;39: 1211–33. 21. Mabilat C, Courvalin P. Development of "oligotyping" for characterization and molecular epidemiology of TEM b-lactamases in members of the family Enterobacteriaceae. Antimicrob Agents Chemother 1990;34:2210–6. 22. Araque M, Nieves B, Ruíz O, Dagert M. Caracterización de plásmidos que median la resistencia a múltiples antibióticos en bacterias gramnegativas de origen nosocomial. Enferm Infecc Microbiol Clin 1997;15:299–305. 23. Araque M, Velazco E. In vitro activity of fleroxacin against multiresistant gram-negative bacilli isolated from patients with nosocomial infections. Intensive Care Med 1998;24:839–44. 24. Sirot D, De Champs C, Chanal C, Labia R, Darfeuille-Michaud A, Perroux R, et al. Translocation of antibiotic resistance determinants including and extended-spectrum b-lactamase between conjugative plasmids of Klebsiella pneumoniae and Escherichia coli. Antimicrob Agents Chemother 1991;35:1576–81. 25. Heritage J, Hawkey P, Todd N, Lewis I. Transposition of the gene encoding a TEM-12 extended-spectrum b-lactamase. Antimicrob Agents Chemother 1992; 36:1981–6.
Copyright Elfos Scientiae 2000 The following images related to this document are available:Line drawing images[ba00022a.gif] [ba00022b.gif] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}