 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnología Aplicada 2000;17:105-108 A microELISA for the quantitation of mouse monoclonal IgM in hybridoma culture supernatants Jorge Fraga Nodarse,1 Juan Carlos Vilaseca Méndez,1 1 Instituto de Medicina Tropical "Pedro Kourí". Autopista Novia
del Mediodía Km 6½ No. 251
Received in October, 1999. Accepted for publication in January, 2000. Code Number: ba00025
ABSTRACT
A microELISA has been set up which is able to sensitively quantitate mouse monoclonal IgM in hybridoma culture supernatants in a suitable range of concentrations. This assay uses ion exchange chromatography-purified rabbit anti-mouse IgM polyclonal antibodies, both in coating capture step and conjugated to horseradish peroxidase in revealing step. It was possible to detect concanavalin A-Sepharose affinity-purified and gel filtration-highly purified mouse monoclonal IgM in the range 190 ng/mL to 12360 ng/mL (sensitivity 125.5 ng/mL). This assay showed a variation coefficient of 4.50% for intraplate repeatability and 7.50% for interplate reproducibility. When compared with an alternative ELISA test that uses a commercial goat anti-IgM-alkaline phosphatase conjugate a correlation coefficient r = 0.92 was found. With this assay, the quality control program of the authors. hybridoma unit is completed, which was developed with their own resources. Keywords: ELISA, IgM, monoclonal antibody, quantitation RESUMEN MicroELISA para la cuantificación de IgM monoclonal de ratón en el sobrenadante de cultivos de hibridomas. Se optimizó un ensayo ELISA que es capaz de cuantificar IgM monoclonal de ratón con gran sensibilidad en el sobrenadante de cultivos de hibridomas en un rango apropiado de concentraciones. Este ensayo utiliza anticuerpos policlonales anti-IgM de ratón generados en conejo y purificados por cromatografía de intercambio iónico. Estos anticuerpos son utilizados como recubrimiento y conjugados con peroxidasa de rábano picante. El ensayo detectó IgM monoclonal de ratón purificada mediante cromatografía de afinidad con concanavalina A-Sefarosa, y mediante filtración en gel, en un rango de cuantificación de 190 ng/mL a 12360 ng/mL (sensibilidad 125,5 ng/mL). Este ensayo mostró repetibilidad intraplaca con un coeficiente de variación (CV) de 4,50% y una reproducibilidad interplaca con un CV de 7,50%. Cuando se comparó con un sistema similar que utilizó el conjugado comercial anti-IgM de ratón-fosfatasa alcalina, se obtuvo un coeficiente r = 0,92. Con este ensayo, se completa un programa de control de la calidad de la unidad de hibridomas de los autores de este trabajo, desarrollada con sus propios recursos. Palabras claves: anticuerpo monoclonal, cuantificación, ELISA, IgM
Introduction
The technology for generation and production of monoclonal antibodies (MAb) has become a basic element in the fields of biomedicine and biotechnology, due to the enormous possibilities of application since its original report in 1975 [1]. Quality control of both MAb generation and production is based, among others, on the accurate quantitation of the molecule in order to estimate the productive capability of a hybridoma and the availability of antibody in a particular production time. On the other hand, such a determination is part of the current characterization of MAbs [2]. Quantitation is critical in the production process using large-scale biorreactors for human-injectable preparations, in which it is important to follow up the pharmacokinetics of the molecule [3, 4]. Up to now, cancer applications have used IgG as "the magic bullet" because there is a greater molecular accessibility to reach and penetrate tumors. It is reasonable to expect that in relation to infectious diseases, the voluminous pentameric IgM molecule would have a better performance for its theoretical ability to agglutinate microbes (bacteria, yeast, large viruses) and protozoa in vivo [5]. Since its original description in 1971, the enzyme-linked immunosorbent assay (ELISA) has become a formidable analytical and practical tool for detecting, monitoring and diagnosing several antibody-detectable analytes due to its high specificity and sensitivity, and due to the large number of samples that can be automatically processed [6]. For that reason, ELISA has been used more frequently for quantitating immunoglobulins, compared with other methods as total protein determination [7], radial immunodiffusion [8, 9], immunofluorescence [10], and agglutination on microplates [11]. Some ELISA assays have been reported for quantitating monoclonal immunoglobulins in biological fluids and mainly in hybridoma supernatants, but the "sandwich" type is currently the most used [12]. Commonly, purified antibodies are used as standard analytes for constructing the quantitating curve [13]. On the other hand, it is not recommendable to use MAbs as coating molecule (breaking not only deletional but dominant tolerance) in order to include an anti-"species" monoclonal conjugate avoiding the overall recognition (by-passing the analyte). MAbs recognize a single epitope with uniform specificity and affinity, whereas polyclonal antibodies recognize several different epitopes on a given antigen and each epitope with a variable affinity. Indeed, the lower affinity of MAb would account for their frequently lower detection levels in ELISA [14]. Another problem using MAbs as capture antibodies in ELISA compared to polyclonal antibodies is that some MAbs have a poor performance once they are coating plastic [15]. In this sense, polyclonal anti-"species" (mainly mouse) antibodies are highly recommended for coating step [12, 16, 17]. However, a lower sensitivity is obtained when polyclonal antibodies are used in capture and detection steps [17]. With the development of rat MAbs by Bazin in 1982 [18], a higher sensitivity in the assay was obtained. To this aim, Delaunay et al. in 1990 [16] used rat anti-kappa light chain as coating and detecting antibody in an ELISA for mouse monoclonal IgM quantitation in hybridoma supernatants. Currently, there are no quantitative mouse IgM ELISA kits available in the market. It is necessary to purchase independent antibodies and to set up assays regarding quality control programs. In this paper, a sandwich microELISA system for quantitating mouse IgM MAbs in hybridoma culture supernatants is reported, which is completely independent of the idiotypic specificity. This assay uses purified monoclonal mouse IgM as standard, and purified rabbit anti-mouse IgM polyclonal antibodies as coating and conjugated antibodies in the ELISA.
Materials and Methods
Antibodies
Two-step purification of mouse IgM MAb. Ascitic fluid was applied to a Concanavalin A-Sepharose column (Amersham Pharmacia Biotech, Sweden) and eluted with alpha-methyl-mannoside. The IgM fraction was then applied isocratically to a gel filtration Superdex 200 HI Load XK 26/60 system (Amersham Pharmacia Biotech, Sweden) [19]. The purity of the IgM was verified by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) [20], and the identity of the molecule was confirmed by a two-step ELISA consisting in an IgM peak coated to the plate and a commercial anti-mouse IgM-peroxidase. This purified IgM was used for making the standard curve in the quantitative ELISA. Preparation of rabbit anti-mouse polyclonal antibody. Purified IgM was quantitated both spectrophotometrically and by the bicinchoninic acid method [21]. Rabbits were immunized subcutaneously by injection of 100 mg of IgM in complete Freund. s adjuvant (Sigma, St. Louis, USA). Doses were repeated in incomplete Freund. s adjuvant every other week until a titer of 1:32 was obtained, as determined by the double immunodiffusion method [22]. Polyclonal antibodies were purified by 50% ammonium chloride salting-out and ion-exchange chromatography in a prepacked Fast Flow HI-Load Q-Sepharose XK 26/20 column (Amersham Pharmacia Biotech, Sweden) [23]. This preparation was used as coating antibody for IgM quantitation and also conjugated to horseradish peroxidase (HPRO) (P-8375, type VI, Sigma, St Louis, USA) by the sodium peryodate method [24].
"Sandwich" ELISA (s-ELISA) for mouse IgM quantitation with purified antibodies Ninety-six-well flat-bottomed ELISA plates (Maxi Sorp, Nunc, Denmark) were coated with 100 mL of rabbit anti-mouse antibodies at a concentration optimally determined (concentration range of capturing antibody 2. 30 mg/mL), for 2 h at 37 ºC in 50 mmol/L coating buffer (Na2CO3/NaHCO3) pH 9.5. The plate was washed four times with phosphate-buffered saline (PBS, 0.1 M NaCl, 2 mM KCl, 0.01 M KH2PO4) containing 0.05% Tween 20 (PBS-T). Blocking step was performed using 1% casein (E. Merck, Darmstadt, Germany) in PBS (150 mL per well), for 1 h at 37 ºC. Plates were then washed four times with PBS-T. One-hundred microliters of purified IgM to several dilutions (from 25 to 12,360 ng/mL), and mouse hybridoma supernatants to appropriate dilutions (RPMI 1640 metabolized medium, Sigma, St. Louis, USA), were added to separated wells and the plates were kept for 1 h at 37 ºC. After six washes with PBS-T, 100 mL of HPRO-conjugated anti-mouse antibodies were added to an optimized working dilution and the plates were incubated for 1 h at 37 ºC. After washing eight times with PBS-T, the substrate (o-phenylenediamine, E. Merck, Darmstadt, Germany; 0.4 mg/mL in 0.1 mol/L citrate/phosphate buffer pH 5.5, 0.04% H2O2) was added and the reaction was stopped 20 min later by adding 50 mL of 12.5% H2SO4 (v/v). P3A63Ag8.653 plasmocytoma cell-conditioned RPMI medium (ATCC, Rockville, MD, USA) and antidengue virus IgG hybridoma supernatants were used as negative control, and PBS-T was used as zero-standard. Incubation times and washing procedures were performed as previously reported [2].
ELISA for mouse IgM quantitation using a commercial conjugate The "sandwich" ELISA using a commercial conjugate (s-ca-ELISA) was performed as described for s-ELISA, but with some modifications. Briefly, a goat anti-mouse alkaline phosphatase conjugate (Sigma, St. Louis, USA) diluted 1:1000 in PBS-T and foetal bovine serum (FBS) 4% was used, and 1 mg/mL r-nitrophenylphosphate (Sigma, St. Louis, USA) in 0.1 mol/L diethanolamine buffer pH 9.8 was used as substrate. The reaction was stopped 30 min later by adding 50 mL of 0.2 N NaOH.
Setting up conjugate in the s-ELISA
For estimating the best conjugate working dilution (1:500. 1:4000) in the s-ELISA, the ratio of maximal optical density to zero-standard optical density was taken into account in the assay previously described.
Linear regression and sensitivity determination The best fit for the standard curve was performed by a graphical dispersion and regression using Office 97 Microsoft Excel (Microsoft® Corporation, USA). Sensitivity in terms of minimal detectable quantity was estimated also taking into account the accuracy of the zero-standard and the maximal absorbance level, as described by Kurone et al. [25] by the formula:
The OD limit should be introduced in the linearized (logarithmic transformation) standard curve for estimating the minimal antigen concentration. Reproducibility and repeatability were evaluated by determining the variation coefficient. Interplate variation coefficient (reproducibility) was determined by assaying the standard curve points in four assays, and intraplate variation coefficient (repeatability) was determined by assaying the standard curve points four times in a single assay.
Correlation Culture supernatants from different IgM hybridomas were used in both s-ELISA and s-ca-ELISA and the respective concentrations were determined. A correlation coefficient for illustrating such association was calculated using Office 97 Microsoft Excel (Microsoft® Corporation, USA).
Results
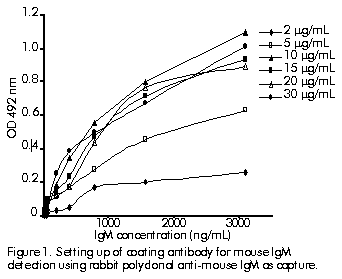
Figure 1 shows the coating antibody setting up for mouse IgM detection using a rabbit polyclonal anti-mouse IgM as capture antibody and an anti-mouse IgM-peroxidase conjugate as detecting antibody. Figure 1. Setting up of coating antibody for mouse IgM detection using rabbit polyclonal anti-mouse IgM as capture. The optimal concentration for the coating antibody was 10 mg/mL. The ratio maximal optical density to zero-standard optical density for different conjugate concentrations, revealed 1:1000 to be the best dilution assayed with a lower zero-standard level (Table).
Table. Ratio of maximal optical density (OD) to zero-standard OD for different conjugate dilutions in s-ELISA.
Standard curve for IgM quantitation using s-ca-ELISA
Figure 2 shows the standard quantitative curve for IgM using a commercial anti-mouse IgM alkaline phosphatase conjugate (s-ca-ELISA). The detection range was adjusted between 190 and 6180 ng/mL and the sensitivity for detection of IgM in hybridoma supernatants was calculated as 373.3 ng/mL. Repeatability was established with a variation coefficient (intraplates) of 4.80% and a reproducibility (interplates) of 7.20%. Figure 2. Standard curve for quantitating mouse IgM monoclonal antibodies by s-ca-ELISA. The appropriate transformed curve is referred.
Standard curve for mouse IgM quantitation using s-ELISA
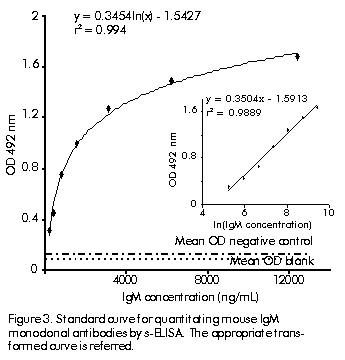
Figure 3 shows the standard quantitative curve for IgM using a rabbit polyclonal anti-mouse IgM as capture antibody and an anti-mouse IgM-peroxidase conjugate as detecting antibody in the s-ELISA. Detection was ranged between 190 and 12,360 ng/mL and the sensitivity for detection of IgM in hybridoma supernatants was estimated in 125.5 ng/mL. Repeatability was established with a variation coefficient (intraplates) of 4.50% and a reproducibility (interplates) of 7.5%. The range (190. 12,360 ng/mL) was selected from a starting point of 25 ng/mL for IgM taking into account reproducibility accuracy criteria in terms of variation coefficient (data not shown). Figure 3. Standard curve for quantitating mouse IgM monoclonal antibodies by s-ELISA. The appropriate transformed curve is referred.
Correlation between s-ELISA and s-ca-ELISA
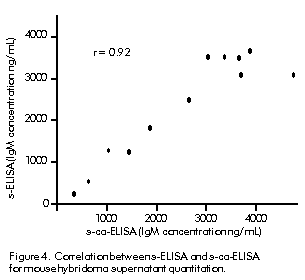
Figure 4 shows point dispersion when quantitative determination was performed by both s-ELISA and s-ca-ELISA. Correlation coefficient was calculated as 0.92. Figure 4. Correlation between s-ELISA and s-ca-ELISA for mouse hybridoma supernatant quantitation.
Discussion
A sandwich ELISA for IgM quantitation has been described, which uses rabbit-purified both coating and conjugated antibodies. In 1990, Delaunay et al. [16] proposed an ELISA assay for mouse monoclonal IgM quantitation in hybridoma supernatants using a rat anti-kappa light chain as coating antibody and conjugated to peroxidase, and using a standard curve based on mouse purified serum. They ranged the assay from 2 to 30 ng/mL, showing a sensitivity of 2 ng/mL with variation coefficients of 3.1% and 4.0% for repeatability and reproducibility, respectively. The assay showed a sensitivity 60-fold lower with a wider quantitation range (between 190 and 12,360 ng/mL). This range is favoring our analytical aim in terms of detecting IgM in the supernatants in the range 1. 20 mg/mL. Using a lower sample dilution in order to have an antibody concentration in the quantitation range of the ELISA procedure, the assay can be performed with a lower dilution error. Note that these authors used rat anti-kappa light chain capturing and detecting antibodies in order to increase the overall sensitivity of the assay. Although 95% of mouse light chain is kappa (k) [26], this approach does not permit quantitation of lambda light chain (l) and consequently the MAbs must be tested for k or l. The loss of sensitivity in the assay that used capturing and detecting polyclonal antibodies was due to the competition for a common epitope between the capture polyclonal antibody and the secondary polyclonal antibody-enzyme conjugate, which causes a steric hindrance [17]. Although in 1986 Butler et al. [17] concluded that, compared to polyclonal antibodies, MAbs usually have not a sufficiently high affinity to permit their use as "immunoglobulin capture and detection" antibodies. Moreover, some MAbs have a poor performance once coated to plastic [15]. Accordingly, Buttler et al. did not recommend MAb anti-immunoglobulins in a sandwich ELISA when a polyclonal alternative was available.
In order to circumvent the loss of sensitivity due to steric hindrance in symmetric assays, Buttler et al. advised the use of an asymmetrical configuration for the measurement of immunoglobulins by sandwich ELISA [17]. Anyway, even with polyclonal purified antibodies, it was possible to obtain values of repeatability and reproducibility below 10% of variation coefficient. In 1989, Klein-Schneegans et al. [8] proposed an ELISA for mouse monoclonal IgM quantitation using rabbit anti-mouse polyclonal IgM as coating antibody and rat monoclonal biotin-conjugated anti-mouse IgM. These authors ranged widely the assay for 1. 1000 ng/mL. Using purified MAbs as standard, the variation coefficient was slightly higher than the 10% recommended by Tijssen in 1985 [14], whereas the variation coefficient was lower than 10% when standard serum was used. The assay proposed in this article shows a variation coefficient lower than 10% when purified MAbs are used as standard and polyclonal serum as capturing antibodies.
Conclusion
A sandwich microELISA for mouse IgM quantitation is presented, which is useful for detecting this isotype of MAb in hybridoma supernatants in the range 190 to 12,360 ng/mL, with a sensitivity of 125.5 ng/mL and an acceptable precision with a variation coefficient below 10%. References 1. Köhler G, Milstein C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975;256: 495–7. 2. Vilaseca JC, Pupo M, Bernal M, Matamoros L, Gordillo S, Rodríguez H, et al. Quantitative ELISA for mouse monoclonal antibody. Determination of culture supernatants and in human serum. Hybridoma 1997;16(6):557–62. 3. Tiny van der Velden AM, Martin JM, Bewey EC. New developments in the cultivation of hybridoma cells in homogeneous continuos perfusion systems. In: Tiny van der Velden AM, Trebout RF, editors. Monoclonal antibodies for therapy, prevention’s and in vivo diagnosis of human disease. Amsterdam: Elsevier; 1990. p.45–54. 4. Trebout RF. Tissue culture in hollow-fibre systems implications for downstream processing and stability analysis. In: Tiny van der Velden AM, Trebout RF, editors. Monoclonal antibodies for therapy, prevention and in vivo diagnosis of human disease. Amsterdam: Elsevier; 1990. p.65–71. 5. Greeman RL, Shein RM, Martin MA, Wendel RP, Intyre NR, Emmanuel G, et al. A controlled clinical trial of E5 murine monoclonal IgM antibody to endotoxin in the treatment of gram-negative sepsis. The XOMA Sepsis Study Group. JAMA 1991; 266(8):1097–102. 6. Voller A, Bidwell DE, Bartlett A, editors. The ELISA: a guide with abstracts of microplate applications. Dynatcch. Europe, Borough. House, Ruedu. Pre. Guernsey, GB; 1979. p.42–50. 7. Manil L, Motte P, Tcroalen F, Bohuon C, Bellet D. Evaluation of protocols for purification’s of mouse monoclonal antibodies. J Immunol Meth 1986;90:25–9. 8. Klein-Schneegans A, Gaveriaux C, Fontaneau P, Loor F. Indirect double sandwich ELISA for the specific and quantitative measurement of mouse IgM, IgA and IgG subclass. J Immunol Meth 1989;119: 117–25. 9. Schmidt PN, Blirup S, Sevendsen PJ, Nandall, JH. Characterization and quantifications of plasma proteins excreted in faeces from healthy humans. Scan J Clin Lab Invest 1995;55(1):35–45. 10. Broeder HW, Groen J, van Steenis G, Osterhaus AD. An enzyme-linked immunosorbent assay for the detection of polyomavirus specific antibodies in laboratory mice. Lab Anim 1994;28(3):257–61. 11. Aubert D, Foundrinier F, Kaltenbach ML, Guyot WD, Marx-Chemla C, Geers R, et al. Automated reading and processing of quantitative IgG, IgM, IgA and IgE isotypic agglutination results in microplates. Development and application in parasitology-mycology. J Immunol Meth 1995;186 (2):323–8. 12. Fleming JO, Pen LB. Measurement of the concentration of murine IgG monoclonal antibody in hybridoma supernants and ascities in absolute units by sensitive and reliable enzyme-linked immunosorbent assays (ELISA). J Immunol Meth 1988;110:11–8. 13. Mushen RE, Guest AR, Scott ML. Quantitation of mononclonal antibodies by ELISA. The use of purified mouse IgG and mouse IgM monoclonal antibodies as standards in a quantitative ELISA measuring monoclonal antibodies procedure by cell culture. J Immunol Meth 1993; 162(1):77–83. 14. Tijssen P, editor. Practice and theory of enzyme immunoassay. Amsterdam: Elsevier; 1985. 15. Kemeny DM, editor. A practical guide to ELISA. Oxford, (UK): Pergammon Press; 1991. 16. Delaunay T, Louhahed J, Bazin H. Rat (and mouse) monoclonal antibodies VIII. ELISA measurement of Ig production in mouse hybridoma culture supernatants. J Immunol Meth 1990;131:33–9. 17. Butler JE, Spradling JE, Suter M, Dierks SE, Heyermann H, Peterman JH. The immunochemistry of sandwich ELISAs I. The binding characteristics of immunoglobulins to monoclonal and polyclonal capture antibodies adsorbed on plastic and their detection by symmetrical and asymmetrical antibody enzyme conjugates. Mol Immunol 1986;23:971–4. 18. Bazin H. Production of rat monoclonal with the LOU rat nonsecreting IR 983F myeloma cell line. In: Peeters H, editor. Protides of the Biological Fluids. Oxford (UK): Pergammon Press; 1982. p.615. 19. Weinstein Y, Givel D, Strausbarch PH. The fractionations of immunoglobulins with insolubilized concanavalin A. J Immunol Meth 1972;109:1402–4. 20. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1974;227: 680–5. 21. Smith PK, Krohn, RI, Hermanson GT, Mallia AK, Gartner FH. Measurement of protein using bicinchoninic acid. Annal Biochem 1985;150(1):76–85. 22. Ouchtherlony O, Nilsson LA. Immunodiffusion and immunoelectrophoresis. In: Weir DM, editor. Handbook of experimental immunology. Oxford Black Well. Sci. Pub. Oxford, London, Edinburg, Melburne; 1978. p.191–4. 23. Karlsson E, Ryden L, Brewer J. Ion exchange chromatography. In: Janson JC, Ryden L, editors. Protein purification: principles, high-resolution methods and applications. New York: VCH; 1989. p.107–15. 24. Nakane PJ, Kawasi KG. Peroxidase-labeled antibody: a new method of conjugation. J Histochem Cystochem 1974; 22(12):1084–91. 25. Kuronen I, Hokko H, Parviainen M. Production of monoclonal and polyclonal antibodies against human osteocalcin sequences and development of two-site ELISA for intact human osteocalcin. J Immunol Meth 1993;263:233–4. 26. Goding JW, editor. Monoclonal antibodies: principle and practice, 2nd ed. London: Academic Press; 1986. p.9. The following images related to this document are available:Line drawing images[ba00025e.gif] [ba00025a.gif] [ba00025c.gif] [ba00025d.gif] [ba00025b.gif] | ||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}