 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnología Aplicada, Volume 17, July-September 2000, pp. 171-176 Aumento del rendimiento de tomate (Lycopersicon esculentum Mill) cuando la raíz se desarrolla colonizada por microorganismos @ Guillermo Carrillo-Castañeda,1 Juana Juárez Muñoz,1 Dionicio Ruiz Landa,2 Rafael Müller García2 1Colegio de Postgraduados. Montecillo, México 56230, México.
E-mail: carrillo@colpos.colpos.mx Recibido en marzo de 1999. Aprobado en mayo de 2000. Code Number: BA00051 RESUMEN Los microorganismos del suelo modifican los patrones de desarrollo y producción de los cultivos a través de mecanismos que se conocen poco y que son mediados por compuestos con actividad biológica. Parcelas de plantas de tomate (Lycopersicon esculentum Mill) inoculadas con aislamientos de Pseudomonas fluorescens, lograron producir hasta 45% más (A9m) que las plantas sin inocular. A los 45 días después del transplante, estas plantas también fueron las de mayor altura. Las plantas inoculadas con el aislamiento A9m —las de mayor producción— iniciaron la floración 6 días antes que las plantas sin inocular. Sin embargo, ninguna de las plantas inoculadas tuvieron mayor volumen radical promedio que las plantas controles, a pesar de que se ha demostrado que las cuatro especies de microorganismos usadas producen ácido indolacético. En estos ensayos, los aislamientos de P. fluorescens promovieron las características indicadas en las plantas mediante expresiones comunes en la especie. Estas expresiones prevalecieron a pesar de la microflora del lugar. Palabras clave: desarrollo vegetal, interacción planta-microorganismo, raíz, rendimiento, tomate ABSTRACT Increase of tomato (Lycopersicon esculentum Mill) yield when the root develops colonized by microorganisms. Soil-borne microorganisms modify the patterns of plant development and crop yields through an array of mechanisms in which sorts of compounds with biological activity play an important role. Plots of tomato plants (Lycopersicon esculentum Mill) inoculated with isolates of Pseudomonas fluorescens (isolate A9m) yielded up to 45% more and bloomed 6 days before, compared to the uninoculated plants. These plants reached the highest lenghts 45 days after potting. None of the inoculated plants showed higher root volumes than the uninoculated plants despite the fact that in these trials we used four species capable of producing indolacetic acid. In these trials, the isolates of P. fluorescens promoted the indicated expressions in the plants through a common expression performed by these microbial species. Their effects prevailed upon the potential effects of the soil-borne microorganisms. Keywords: plant growth, plant-microorganism interaction, root, tomato, yield Introducción De la diversidad de especies de microorganismos que componen la microflora del suelo, muchas producen compuestos con actividad biológica que son capaces de interferir con el desarrollo de microorganismos fitopatógenos [1-9]. En investigaciones realizadas sobre las interacciones entre diversas plantas y ciertos microorganismos promotores del desarrollo vegetal, cuyas siglas en inglés son PGPR, la raíz desempeña un papel central por ser el órgano de la planta que es colonizado en primera instancia [10-12]. En esta interacción se promueve tanto el crecimiento de la raíz [13-17] como la capacidad de absorción de sustancias nutritivas [18]. Ciertas especies de microorganismos de diversos géneros han sido utilizadas en la práctica para incrementar la producción de múltiples cultivos, ya sea por su participación en el control biológico de hongos y bacterias fitopatógenos [9], por su relación con la inducción del estado de resistencia sistémica adquirida [19], o por su capacidad de fijar nitrógeno atmosférico [20, 21], entre una diversidad de actividades biológicas [2, 3, 5, 22]. En especial, Pseudomonas fluorescens es ampliamente conocida por promover el desarrollo de algunos cultivos. Sin embargo, los variados mecanismos mediante los cuales esta acción general se lleva a cabo no son plenamente conocidos y, por lo tanto, es necesario determinar con precisión su efecto en la biología de la planta beneficiada. En el presente trabajo, se determinó el efecto de la inoculación en el desarrollo de la raíz y de la planta, así como en la producción, con la expectativa de utilizar la información generada en estas investigaciones para establecer interacciones de la manera más apropiada y práctica. El propósito final es incrementar significativamente los rendimientos de cultivos importantes. Materiales y Métodos Material biológico Se utilizaron semillas de Lycopersicon esculentum "Río Grande" y "Floradade"; Pseudomonas solanacearum y los aislamientos de microorganismos M96-20 y M9-22; P. fluorescens Avm, A7, A9 y A9m de este laboratorio; Rhizobium leguminosarum variedad phaseoli CPMéx 44 y 46, donados por la Sección de Microbiología del Instituto de Recursos Naturales, Colegio de Postgraduados; Azospirillum lipoferum Cd, Sp7, Sp59, UAP4, UAP40, UAP73 y Acetobacter paspali (Apas), proporcionados en el Departamento de Microbiología de la Universidad Autónoma de Puebla. Medios de cultivo Se empleó el medio YMB [23] en estado sólido (YMBS) suplido con antibióticos (en mg/L): YMBScr (carbenicilina 50), YMBScf (cloranfenicol 30), YMBSkm (kanamicina 50), YMBSrf (rifampicina 30) y YMBStet (tetraciclina 15). También se emplearon el medio de cultivo mínimo RM [24], el medio carente de hierro RM-Fe (medio RM sin FeCl3), y los medios ricos BK [23] y PDA (infusión de papa 200 g/L, glucosa 20 g/L y agar 16 g/L). Estos medios en estado sólido contenían 1,6% (p/v) de agar grado bacteriológico. Inoculación de la semilla Para la inoculación de las semillas de ambas variedades con los microorganismos indicados, se partió de 10 mL de cada cultivo desarrollado en el medio RM-Fe (densidad óptica entre 0,6 y 0,9 determinada a 660 nm en un espectrofotómetro Coleman Jr modelo 6/20, EUA). Después de centrifugar a 15 900 xg por 30 min a 4 ºC, las células fueron resuspendidas en 2,5 mL de solución salina (NaCl 0,85% [p/v]) para finalmente mezclarlas con 30 mg de alginato de sodio. Las semillas fueron encapsuladas con esta suspensión. Efecto de la inoculación de la semilla en la capacidad de germinación Para determinar el efecto de la inoculación en la capacidad de germinación, se colocaron lotes de 100 semillas encapsuladas de la variedad Floradade en placas Petri sobre dos capas de papel filtro húmedo y se conservaron a 28-30 ºC en la oscuridad. Se consideró que la semilla había germinado cuando la raíz quebró la testa. Determinación del efecto de la inoculación en el desarrollo del cultivo y la producción Establecimiento del cultivo en invernadero. Se encapsularon (inocularon) series de cinco lotes de semilla de la variedad Río Grande con cada una de las 12 cepas de microorganismos y se conservaron a temperatura ambiente en frascos con restos de agua destilada. Estas semillas se sembraron en las primeras 24 h en bandejas germinadoras que contenían tierra, lo que permitió el desarrollo de las plántulas en el invernadero hasta que alcanzaron unos 12 cm de altura. Las series de plántulas se transplantaron al suelo para establecer los cultivos de acuerdo a un diseño experimental de bloques al azar con cinco repeticiones. La unidad experimental estaba formada por ocho plántulas colocadas en parcelas de 1,35 m2. En el diseño se incluyó un control sin inocular, también con cinco repeticiones. La parcela útil estaba formada por las seis plantas centrales. Las plantas se fertilizaron a los 15 y 45 días después del transplante, sólo con 50% del nitrógeno de la fórmula de fertilización (180:60:30 N:P:K) que los especialistas recomiendan para el cultivo. El tutoreo de la planta se practicó de forma individual, se eliminaron las hojas dañadas y se efectuó el deshierbe de forma manual en tres ocasiones. Como medida preventiva, se aplicó Ambush y Thionex contra la mosquita blanca (Bemicia tabaci), y Cupravit y Bravo 720 contra el tizón tardío (Phytophtora infestans). La cosecha se llevó a cabo de forma manual y cada fruto fue pesado una vez que alcanzó la madurez comercial. Las características evaluadas fueron: altura de la planta, determinada a los 45 días después del transplante; días antes de la floración; volumen radical y rendimiento. En cada caso se llevó a cabo el análisis de varianza de acuerdo al diseño experimental utilizado y se hicieron las comparaciones de medias de Tukey (a = 0,05). Determinación de la capacidad de los cultivos microbianos para desarrollarse en los medios RM-Fe y BK La capacidad de desarrollo de los cultivos A7, A9, A9m y Avm se determinó mediante la inoculación de 25 mL de medio RM-Fe con 0,1 mL de suspensión bacteriana de cultivos desarrollados en medio BK. Posteriormente, se incubó con agitación a 28-30 ºC durante 36 h. La densidad óptica se determinó a 660 nm en un espectrofotómetro (Coleman Jr modelo 6/20, EUA). Determinación de la capacidad de los microorganismos promotores del desarrollo vegetal para producir sideróforos e interferir con el desarrollo de otros microorganismos La capacidad de producción de sideróforos se evaluó mediante el método universal de Schwyn y Neilands [25], y la capacidad de interferir con el desarrollo de P. solanacearum —microorganismo utilizado para esta prueba microbiológica en este laboratorio— se definió mediante la aparición de halos de inhibición al cultivarlos simultáneamente en los medios de cultivo indicados. Resistencia a antibióticos El patrón de resistencia a antibióticos de cada uno de los cultivos se determinó a partir del estriado de las células bacterianas en los medios con antibiótico y su posterior incubación a 28-30 ºC durante 72 h. Determinación de las relaciones filogenéticas entre las cepas de P. fluorescens Las relaciones filogenéticas de los aislamientos indicados se determinaron sobre la base de los patrones de fragmentos de ADN obtenidos por amplificación (reacción en cadena de la polimerasa, RCP) en un termociclador MJ Research modelo PTC 100 (MJ Research Inc., EUA) con el siguiente programa: un ciclo inicial de desnaturalización a 94 ºC durante 1 min y 38 ciclos de amplificación (desnaturalización a 94 ºC por 30 s los tres primeros ciclos y por 15 s el resto, hibridación a 35 ºC por 30 s y polimerización a 72 ºC por 90 s). Se utilizaron los oligonucleótidos iniciadores que se muestran en la Tabla 1. El ADN que se utilizó como molde se obtuvo mediante lisis alcalina. El grado de similitud de los aislamientos se evaluó a partir de los patrones de fragmentos obtenidos con cuatro iniciadores (Tabla 1), y se registraron las bandas como presentes (1) o ausentes (0) en una matriz muestra x banda. Los dendrogramas de las relaciones filogenéticas se construyeron basados en la matriz de similitud, mediante el uso del coeficiente de Jaccard y con la aplicación del análisis de agrupamiento UPGMA y el programa de computación NTSYS-PC versión 1.8 [26]. Tabla 1. Lista de oligonucleótidos usados para la amplificación de ADN.
*Oligonucleótidos usados para el análisis de datos. Resultados El objetivo de este trabajo ha sido caracterizar los aislamientos en estudio a partir de tres tipos de criterios: 1) en asociación, mediante la determinación de la capacidad para promover el desarrollo de la planta desde la germinación de la semilla hasta la etapa de producción; 2) en forma aislada determinando la capacidad de desarrollo en un medio deficiente y otro rico, la producción de sideróforos y la habilidad de interferir con el desarrollo de microorganismos; y 3) el polimorfismo de fragmentos de ADN y las relaciones filogenéticas. Efecto de los microorganismos en la capacidad de germinación de las semillas de tomate variedad Floradade En primera instancia, se trató de caracterizar los aislamientos o cepas microbianas en estudio sobre la base de los efectos promotores de la germinación de la semilla de tomate, para lo que se requirió utilizar semillas de la variedad Floradade cuya capacidad de germinación, por envejecimiento natural, no rebasa 70%. Los resultados indicaron una disminución de la germinación con el aislamiento A9, mientras que el aislamiento A7 mostró una cinética de germinación diferente, aunque con la excepción del primer caso la capacidad de germinación de la semilla inoculada fue similar a la del control (Tabla 2). Tabla 2. Evaluación de la germinación de lotes de 100 semillas de L. esculentum de la variedad Floradade inoculadas con los aislamientos bacterianos. Las células fueron mezcladas con el alginato usado para encapsular la semilla. Las semillas fueron incubadas a 26 ± 2 ºC con fotoperíodo de 16 h de luz.
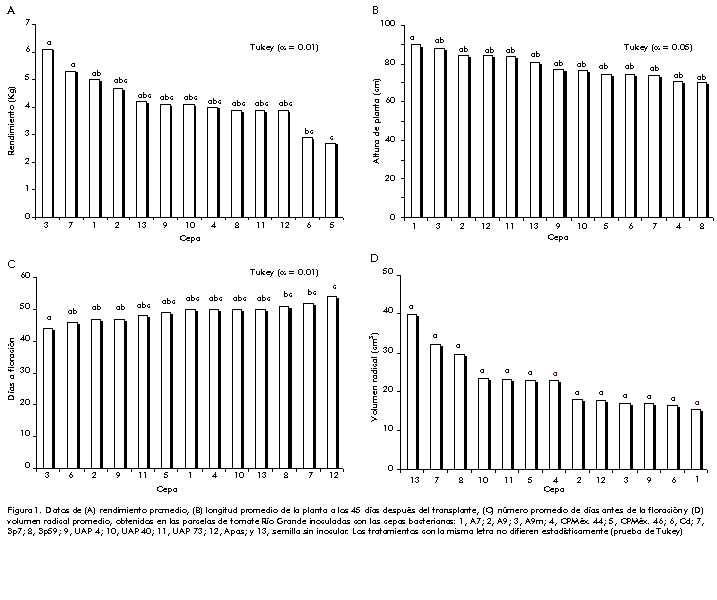
Efecto de la inoculación de la semilla de la variedad Río Grande en características agronómicas importantes y en la producción de tomate Las características estudiadas, en especial la precocidad y el rendimiento, son parámetros importantes en los procesos de producción agrícola. En los resultados que se presentan en la Figura 1, llama la atención que en las parcelas de plantas inoculadas con el aislamiento A9m se obtuvo 45% más de producción, puesto que se cosechó, como promedio, 6 kg por parcela útil, en comparación con la producción de las plantas que no fueron inoculadas y cuya producción promedio alcanzó 4,2 kg (Figura 1A). Las plantas de menor producción fueron las inoculadas con el aislamiento CPMéx 46 (2,7 kg). Para este carácter, se encontraron diferencias altamente significativas por el efecto de la inoculación, según la prueba de Tukey (a = 0,01) (Tabla 3). Figura 1. Datos de (A) rendimiento promedio, (B) longitud promedio de la planta a los 45 días después del transplante, (C) número promedio de días antes de la floración y (D) volumen radical promedio, obtenidos en las parcelas de tomate Río Grande inoculadas con las cepas bacterianas: 1, A7; 2, A9; 3, A9m; 4, CPMéx. 44; 5, CPMéx. 46; 6, Cd; 7, Sp7; 8, Sp59; 9, UAP 4; 10, UAP 40; 11, UAP 73; 12, Apas; y 13, semilla sin inocular. Los tratamientos con la misma letra no difieren estadísticamente (prueba de Tukey) Tabla 3. Análisis de varianza de los caracteres indicados.
*Significativo con a £ 0,05; **significativo con a £ 0,01. Fueron interesantes los resultados de la longitud de la planta (Figura 1B), la cual se determinó prácticamente al inicio de la floración. Los aislamientos A7, A9m, A9 y Apas promovieron, en ese orden, el máximo desarrollo de las plantas. Las plantas inoculadas con el aislamiento A7 alcanzaron 90 cm de altura como promedio y las que no fueron inoculadas, 81 cm. La promoción del crecimiento de las plantas es un aspecto característico de muchas especies PGPR [14, 27]. Nuevamente, las diferencias que se encontraron para este tratamiento resultaron significativas según Tukey (a = 0,05) (Tabla 3). De manera coincidente, las plantas de mayor producción inoculadas con A9m fueron la más precoces en la floración, proceso que comenzó 6 días antes que en las plantas sin inocular. La floración de las plantas sin inocular se observó a los 50 días. Las plantas inoculadas con A9 iniciaron la floración a los 47 días, las inoculadas con A7 florecieron al mismo tiempo que las plantas controles, y las más tardías fueron las inoculadas con el aislamiento Apas (Figura 1C). Para este tratamiento, las diferencias fueron altamente significativas según Tukey (a = 0,01) (Tabla 3). Los datos de volumen radical (Figura 1D) indican que todas las plantas inoculadas tuvieron, como promedio, valores menores en relación con las plantas control. Las plantas inoculadas con el aislamiento A7 —tercer lugar en rendimiento— presentaron el menor volumen radical (15,4 cm3), y para las inoculadas con el aislamiento A9m —primer lugar en rendimiento— el volumen radical fue de 17 cm3. El máximo coeficiente de variación (55,734%) se detectó para este rasgo; sin embargo, en el análisis efectuado según Tukey no se encontraron diferencias en el volumen radical por el efecto de los tratamientos (Tabla 3). En general, las parcelas inoculadas con los aislamientos de P. fluorescens tenían las plantas de mayor producción, las de mayor altura y las más precoces en cuanto a floración, pero, por el contrario, presentaron volúmenes radicales relativamente reducidos. Basado en la importancia práctica potencial de los resultados obtenidos con los aislamientos de P. fluorescens, se optó por estudiar estos aislamientos e incluir el aislamiento Avm en cada experimento, el cual ha sido estudiado en este laboratorio. Determinación de los rasgos de desarrollo y las expresiones de las cepas de microorganismos seleccionados Los aislamientos estudiados tuvieron la capacidad de desarrollarse en el medio RM-Fe y se detectaron valores de densidad óptica entre 0,46 y 0,6 a las 36 h de cultivo, mientras que los valores determinados en el medio BK fueron 0,76; 0,91; 0,75 y 0,75 para los cultivos A7, A9, A9m y Avm, respectivamente. En períodos de incubación de 48 h, se registraron valores mínimos de densidad óptica de 0,6 cuando se cultivaron en el medio RM-Fe. Estos resultados reflejan la plasticidad de estos microorganismos para desarrollarse, tanto en el medio rico como en el medio deficiente. La prueba química universal para detectar la presencia de sideróforos dio resultados positivos. Esta prueba es específica para comprobar que estos microorganismos son capaces de inducir un sistema de asimilación de hierro de alta afinidad —estrategia propia de muchos microorganismos para sobrevivir en medios carentes de hierro—, y de interferir con el desarrollo del fitopatógeno P. solanacearum. La aparición de halos anaranjados en torno al sitio donde crecen las células de los aislamientos Avm, A7, A9 y A9m (Figura 2A), se debe a la presencia de sistemas de alta afinidad de asimilación de hierro que se expresan en respuesta a la carencia de hierro en el medio RM-Fe. Las células del aislamiento M96-20 que están colocadas en diagonal sobre el medio de cultivo, y que previamente fueron caracterizadas por su incapacidad de producir sideróforos en el laboratorio, no mostraron cambio de color. Esta prueba es selectiva para identificar la actividad de sideróforos. Mediante la determinación de interferencia microbiana, también se demostró la presencia de los halos de inhibición que producen los aislamientos A9m y Avm en el desarrollo de P. solanacearum, microorganismo que es utilizado rutinariamente en nuestro laboratorio para determinar interferencia microbiana. Estos halos de inhibición no sólo se observaron en el medio RM-Fe (Figura 2C), sino también en los medios RM (Figura 2B) y PDA (Figura 2D). Estos resultados se pueden explicar a partir del hecho de que la interferencia microbiana no solamente se produce por la competencia por nutrientes, sino también a través de mecanismos como la producción de antibióticos, según se ha demostrado claramente en el caso de P. fluorescens. Se utilizaron como control los aislamientos M96-20 y M96-22, que son capaces de interferir con el desarrollo de P. solanacearum y están en estudio en nuestro laboratorio en estos momentos. Figura 2. Aspecto de los halos desarrollados. A) Color anaranjado que se obtiene en respuesta a la producción de sideróforos. Los tres aislamientos colocados en diagonal no producen sideróforos. B, C y D) Inhibición del desarrollo de P. solanacearum por cepas de microorganismos PGPR en los medios indicados. (Vea imagen a color en la página 214). Identificación de los aislamientos a partir de los patrones de resistencia a antibióticos Una práctica sencilla para identificar un microorganismo determinado con cierto grado de seguridad, es determinar su patrón de resistencia a varios antibióticos. De los 4 aislamientos que se presentan en la Tabla 4, en el medio con carbenicilina sólo se detectaron las cepas Avm, A7 y A9m, y con un desarrollo lento. De esta forma se pudo diferenciar el aislamiento A9 de los otros tres. Tabla 4. Evaluación del crecimiento de las cepas bacterianas estriadas en medio BK sólido suplido con antibióticos. La determinación se hizo después de 48 h de incubación a 30 ºC.
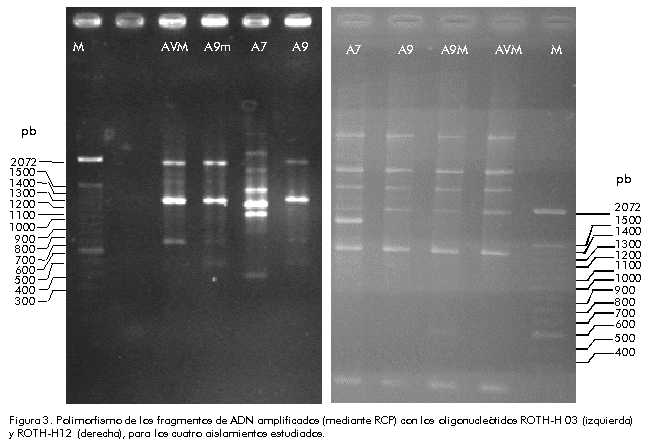
± Crecimiento lento, + crecimiento rápido. Caracterización de los aislamientos microbianos en relación con el polimorfismo de fragmentos amplificados de ADN La otra forma empleada para identificar estos microorganismos, fue el análisis de los patrones de fragmentos de ADN obtenidos mediante la RCP. Sólo con los oligonucleótidos ROTH-H03 y H12 se logró detectar patrones de fragmentos polimórficos específicos del aislamiento A7, lo que permitió diferenciarlo plenamente de los otros tres. Por esa razón, en la Figura 3 sólo se muestran estos dos resultados. Figura 3. Polimorfismo de los fragmentos de ADN amplificados (mediante RCP) con los oligonucleótidos ROTH-H 03 (izquierda) y ROTH-H12 (derecha), para los cuatro aislamientos estudiados. Identificación de las relaciones filogenéticas existentes entre las cepas de P. fluorescens El análisis de los patrones obtenidos con los iniciadores ROTH-G01, ROTH-G02, ROTH-G15, ROTH-H03, ROTH-H09 y ROTH-H12, demostró que los patrones desarrollados con los iniciadores ROTH-G01 y ROTH-G15 no tuvieron la resolución adecuada, por lo que no se incluyeron en el análisis. Con los patrones de los fragmentos de ADN amplificados con los otros 4 oligonucleótidos, se establecieron las relaciones genotípicas existentes entre los microorganismos estudiados. El dendrograma que se muestra en la Figura 4 indica claramente que el organismo menos emparentado es el correspondiente al aislamiento A7. Figura 4. Dendrograma que representa las relaciones filogenéticas entre las cepas indicadas. Discusión Los rendimientos obtenidos para las plantas de tomate inoculadas, están relacionados con las condiciones de cultivo impuestas. Su impacto global está resumido con precisión en los datos de rendimiento de las plantas que no fueron inoculadas. Los niveles bajos de producción de los cultivos son el resultado de la práctica agrícola en tierras poco fértiles o contaminadas de microorganismos fitopatógenos. Estas condiciones de cultivo son precisamente las que permiten que se manifiesten ciertas expresiones de los microorganismos promotores del desarrollo vegetal. Las condiciones óptimas de cultivo pueden invalidar los efectos beneficiosos de estas interacciones. Por lo tanto, para capitalizar al máximo el beneficio de estas interacciones, es necesario determinar, en la práctica y con precisión, hasta qué punto se pueden sustituir ciertos requerimientos del cultivo mediante la inoculación de la semilla con microorganismos seleccionados. Gagné y colaboradores [22] observaron que no se obtenían incrementos significativos en el rendimiento cuando la planta de tomate se inoculaba con microorganismos promotores del desarrollo vegetal en condiciones ambientales óptimas, con luz natural pero a bajas temperaturas que no eran favorables. P. fluorescens es conocido ampliamente por su capacidad de promover el desarrollo de las plantas. Sin embargo, los mecanismos involucrados en este proceso no se conocen plenamente. Para que se produzca este efecto, deben desempeñar un papel importante la eficiencia de la célula bacteriana para colonizar los sitios de crecimiento de la raíz —por los que compiten también los microorganismos nativos— y su capacidad para adaptarse a las condiciones nutricionales del medio ambiente. En este trabajo, se ha demostrado que nuestros aislamientos de P. fluorescens tienen la capacidad de desarrollarse en medios deficientes en nutrientes, lo cual hace que su capacidad para sobrevivir en el ambiente natural sea mayor [4]. Posteriormente, participan otras expresiones mediante las cuales los microorganismos promotores del desarrollo vegetal facilitan la asimilación de elementos nutritivos por la planta [28, 29], promueven el establecimiento de micorrizas [18], desempeñan un papel de biocontrol [30, 31] mediante la inducción de resistencia sistémica adquirida [32-34], o promueven el desarrollo de la planta. Las expresiones relacionadas con el biocontrol o la asimilación de hierro que se observaron para estos microorganismos, podrían explicar los resultados obtenidos al menos en parte. Estas funciones están relacionadas directamente con la concentración del compuesto activo y pueden llevarse a cabo durante la interacción, independientemente de los cambios observados en el volumen radical de las plantas. En otras palabras, los beneficios obtenidos de estas interacciones no fueron mediados por la acción de reguladores del crecimiento vegetal, por lo menos a nivel del sistema radical, puesto que el volumen radical disminuyó en las plantas inoculadas con P. fluorescens que tuvieron los mayores rendimientos. Por lo tanto, consideramos que el desarrollo radical deficiente de estas plantas por el efecto de la inoculación, ha sido compensado por otra u otras actividades de los microorganismos que se tradujeron en incrementos de los rendimientos del cultivo. Se debe tener en cuenta 1) que la actividad biológica de los reguladores del crecimiento vegetal está en función de las concentraciones y no depende únicamente de su presencia o no en el medio; 2) que un microorganismo puede afectar potencialmente a la planta de diferentes formas; y 3) que las condiciones en que se desarrolla el cultivo pueden invalidar o resaltar, o cambiar, el sentido de estos efectos. Una cepa de P. fluorescens productora de ácido indolacético incrementó en 57% el sistema radical en un cultivo, pero provocó una disminución de 45% en otro. Estos autores plantean que una explicación de estos resultados pudiera estar en el grado diferente de sensibilidad de las plantas al ácido indolacético producido por el microorganismo [16]. Loper y Schrot [35] obtuvieron resultados similares con plántulas inoculadas de Beta vulgaris. Es evidente que los aislamientos estudiados en este trabajo han logrado afectar a la planta mediante expresiones que son comunes en la especie, independientemente de sus relaciones filogenéticas. Se debe estudiar con mayor atención el repertorio de compuestos producidos por los microorganismos [36] potencialmente útiles para la agricultura. Se debe tener presente que, además de demostrar la presencia de compuestos producidos por estos microorganismos en condiciones de laboratorio, la concentración de estos compuestos durante la interacción tiene una relación estrecha con los efectos positivos o negativos que causan en la planta (incremento o disminución del desarrollo de un órgano). De la misma forma, se debe tener en cuenta que un microorganismo puede afectar a la planta mediante diversos mecanismos, y que las condiciones de cultivo pueden invalidar o resaltar los efectos de las interacciones. Agradecimientos Los autores agradecen el apoyo económico del CONACYT (0453P-N), que hizo posible el desarrollo de la investigación. Referencias 1. Ahl P, Voisard C, Défago G. Iron bound-siderophores, cyanic acid, and antibiotics involved in suppression of Thielaviopsis basicola by a Pseudomonas fluorescens strain. J Phytopathology 1986;116:121–34. 2. Rodríguez F, Pfender WF. Antibiosis and antagonism of Sclerotinia homoeocarpa and Drechslera poae by Pseudomonas fluorescens Pf-5 in vitro and in planta. Phytopatology 1997;87:614–21. 3. Vanneste JL, Yu J, Bonn WG. Biological control of fire blight using Erwinia herbicola EH252 and Pseudomonas fluorescens A506 separately or in combination. Horticulturae 1996;411:351–4. 4. Carrillo-Castañeda G, Vázquez MRG. Comparative study of siderophore-like activity of Rhizobium phaseoli and Pseudomonas fluorescens. J Plant Nutr 1992; 15:579–90. 5. Nautiyal CS. Selection of chickpea-rhizosphere-competent Pseudomonas fluorescens NBRI1303 antagonistic to Fusarium oxysporum f. sp. ciceris, Rhizoctonia bataticola and Pythium sp. Curr Microbiol 1997;35:52–8. 6. Brisbane PG, Harris JR, Moen R. Inibition of fungi from wheat roots by Pseudomonas fluorescens 2-79 and fungicides. Soil Biol Biochem 1989;21:1019–25. 7. Daniel J, Sullivan O, O’Gara F. Traits of fluorescent Pseudomonas spp. involved in suppression of plant root pathogens. Microbiol Rev 1992;56:662–76. 8. Vanneste JL, Yu J, Bonn WG. Biological control of fire blight using Erwinia herbicola EH252 and Pseudomonas fluorescens A506 separately or in combination. Horticulturae 1996;411:351–4. 9. Neilands JB, Leong SA. Siderophores in relation to plant growth and disease. Ann Rev Plant Physiol 1986;37:187–208. 10. Chiarine L, Bevivino A, Yabacchioni S, Dalmastri C. Inoculation of Burkholderia cepacia. Pseudomonas fluorescens and Enterobacter sp. on Sorghum bicolor: root colonization and plant growth promotion of dual strain inocula. Soil Biol Biochem 1998;30:81–7. 11. Jiang HY, Sato K. Interrelationships between bacterial populations on the root surface of wheat and growth of plant. Soil Sci Plant Nutr 1994;40:683–9. 12. Schroth MN, Hancock JG. Disease-suppressive soil and root-colonizing bacteria. Science 1982;216:1376–81. 13. Germida JJ, Walley FL. Plant growth-promoting rhizobacteria alter rooting patterns and arbuscular mycorrhizal fungi colonization of field-grown sprint wheat. Biol Fert Soil 1997;23:113–20. 14. Fages J, Arsac JF. Sunflower inoculation with Azospirillum and other plant growth promoting rhizobacteria. Plant Soil 1991; 137:87–90. 15. Zaady E, Perevolotsky A, Okon Y. Promotion of plant growth by inoculum with aggregated and single cell suspensions of Azospirillum brasilense Cd. Soil Biol Biochem 1993;25:819–23. 16. Dubeikovsky AN, Mordukhova EA, Kochetkov VV, Polikarpova FY, Boronin AM. Growth promotion of blackcurrant softwood cuttings by recombinant strain Pseudomonas fluorescens BSP53a synthesizing an increased amount of indoleacetic acid. Soil Biol Biochim 1993; 25:1277–81. 17. Rademacher W. Gibberellin formation in microorganisms. Plant Growth Reg 1994;15:303–14. 18. Garbaye J. Helper bacteria: a new dimension to the mycorrhizal symbiosis. New Phytol 1994;128:197–210. 19. Raupach GS, Liu L, Murphy JF, Tuzun S, Kloepper JW. Induced systemic resistance in cucumber and tomato against cucumber mosaic cucumovirus using plant growth-promoting rhizobacteria (PGPR). Plant Dis 1996;80:891–4. 20. Malik KA, Bilal R, Mehnaz S, Rasul G, Mirza MS, Ali S. Association of nitrogen-fixing plant growth promoting rhizobacteria (PGPR) with kallar grass and rice. Plant Soil 1997;194:37–44. 21. Zhang F, Narjes D, Hynes RK, Smith DL. Plant growth promoting rhizobacteria and soybean (Glycine max [L.] Merr.) nodulation and nitrogen fixation at suboptimal root zone temperatures. Ann Bot 1996; 77:453–9. 22. Gagné S, Dehbi L, Le Quéré D, Cayer F, Morin J-L, Lemay R, et al. Increase of greenhouse tomato fruit yields by plant growth-promoting rhizobacteria (PGPR) inoculated into the peat-based growing media. Soil Biol Biochem 1993;25:269–72. 23. Vincent JM. Manual práctico de rizobiología. Hemisferio Sur, Argentina; 1975. 24. Carrillo-Castañeda G, Peralta JRV. Siderophore-like activities in Rhizobium phaseoli. J Plant Nut 1988;11:935–44. 25. Schwyn B, Neilands JB. Universal chemical assay for the detection and determination of siderophores. Anal Biochem 1987;160:47–56. 26. Rohlf FJ. NTSYS-pc numerical taxonomy and multivariate analysis system, version 1.80. Applied Biostatistics Inc, New York; 1993. 27. Frommel MI, Nowak J, Lazarovits G. Growth enhancement and developmental modifications of in vitro grown potato (Solanum tuberosum ssp. tuberosum) as affected by a nonfluorescent Pseudomonas sp. Plant Physiol 1991;96:928–36. 28. Kavimandan SK, Gaur AC. Effect of seed inoculation with Pseudomonas sp. on phosphate uptake and yield of maize. Curr Sci 1971;40:439–40. 29. Duijff BJ, Bakker PAHM, Schippers B. Ferric pseudobactin 358 as iron source for carnation. J Plant Nutr 1994;17:2069–78. 30. Dune C, Delany I, Fenton A, O’Gara F. Mechanisms involved in biocontrol by microbial inoculants. Agronomie 1997; 16:71–9. 31. Mavrodi DV, Ksenzenko VN, Bonsall RF, Cook RJ, Boronin AM, Thomashow LS. A seven-gene locus for synthesis of phenazine-1-carboxylic acid by Pseudomo-nas fluorescens 2-79. J Bacteriol 1998; 180:2541–8. 32. Mercado-Blanco J, Olsson PE, van der Drift KMGM, Thomas-Oates JE, van Loon LC, Bakker PAHM.Analysis of a gene cluster involved in the production of salicylic acid and a new siderophore in Pseudomonas fluorescens WCS374. In: Ogoshi A, editor. Plant growth-promoting rhizobacteria: present status and future prospects. Sapporo: Nakanishi; 1997. p.355–7. 33. Yalpani N, Raskin I. Salicylic acid: a systemic signal in induced plant disease resistance. Trends Microbiol 1993;1:88–91. 34. Wei G, Kloepper JW, Tuzun S. Induced systemic resistance to cucumber diseases and increased plant growth by plant growth-promoting rhizobacteria under field conditions. Phytopathology 1996; 86:221–4. 35. Loper JE, Schrot MN. Influence of bacterial sources of indole-3-acetic acid on root elongation of sugar beet. Phytopathology 1996;76:386–9. 36. Gutiérrez-Manero FJ, Acero N, Lucas JA, Probanza A. The influence of native rhizobacteria on European alder (Alnus glutinosa [L.] Gaertn.) growth. II. Characterisation and biological assays of metabolites from growth promoting and growth inhibiting bacteria. Plant Soil 1996;182:67–74. Copyright Elfos Scientiae 2000 The following images related to this document are available:Photo images[ba00051d.jpg] [ba00051c.jpg] [ba00051b.jpg] [ba00051a.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}