 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnología Aplicada, Volume 17, July-September 2000, pp. 177-182 Isolation and Structure-functional Characterization of Human Colostral Lactoferrin @ Antonio Aguila La O,1 Ana
G Herrera Puerta,1 William Toledo Velázquez,1 1Group for Research on Lactoferrin, Iron Metabolism, and Immunomodulation.
Finlay Institute. Ave. 27. No. 19805, La Lisa. AP 16017, CP 11600, Havana, Cuba.
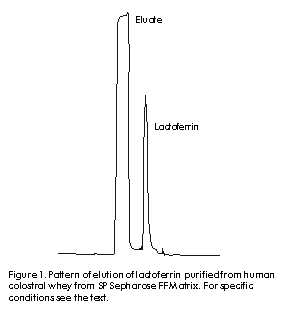
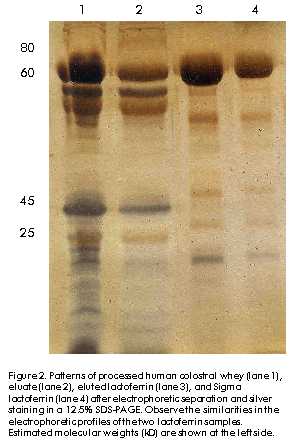
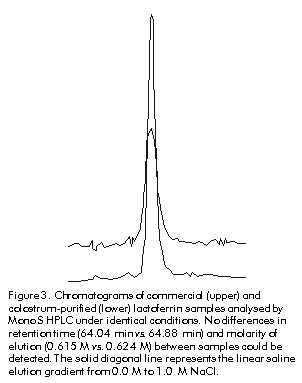
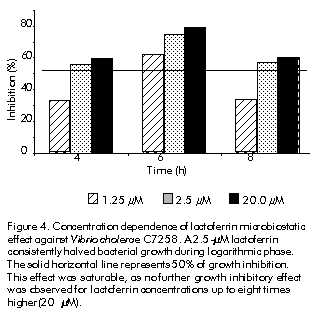
Phone: (53-7) 21 3480; Fax: (53-7) 28 6075; E-mail: aaguila@finlay.edu.cu Received in June, 2000. Accepted for publication in June, 2000. Code Number: BA00052 Abstract Human lactoferrin has been the focus of extensive research because of its antimicrobial, ferrochelating, immunomodulatory, and diagnostic potentialities. To meet the need for homogenous lactoferrin preparations, here we report simple procedures for colostrum processing, chromatographic isolation, and quality control techniques for the evaluation of the most relevant structure-functional properties of lactoferrin. Purity and identity were determined by combination of electrophoretic, densitometric, and immunochemical techniques. Structural integrity was assayed by high-performance liquid chromatography (HPLC), and iron-chelating properties were evaluated by spectroscopic analysis of lactoferrin derivatives. Antimicrobial activity was tested in vitro against Vibrio cholerae C7258, a regional epidemic strain. These procedures allowed the isolation and characterization of substantially pure (98–99%), structurally intact (92–93%), non-toxic, lipopolysaccharide (LPS)-free (less than 0.40 EU/mg of protein), low-iron saturated (below 7% saturation), biologically active (2.50 µM lactoferrin inhibited V. cholerae growth in 50%), and consistently homogenous preparations. The quality of this lactoferrin preparation allows its use in basic studies of the mechanisms of microbial virulence, and in the development of new tools for the diagnosis of gastrointestinal inflammation which are currently being performed by our group. Keywords: antibacterial activity, human colostrum, iron, lactoferrin, LPS, protein purification, Vibrio cholerae Resumen Purificación y caracterización estructural y funcional de la lactoferrina de calostro humano. La lactoferrina humana ha sido intensamente investigada por su potencial antimicrobiano, ferroquelante, inmunomodulador y para el diagnóstico. Dada la necesidad de preparaciones homogéneas de lactoferrina, en este trabajo se reportan procedimientos simples para el procesamiento del calostro y su aislamiento cromatográfico, y técnicas de control de calidad de sus propiedades estructurales y funcionales. La pureza e identidad fueron determinadas mediante técnicas electroforéticas, densitométricas e inmunoquímicas. La integridad estructural se evaluó por cromatografía líquida de alta presión (HPLC), y las propiedades ferroquelantes se evaluaron por análisis espectroscópico de sus derivados. La actividad antimicrobiana se evaluó in vitro contra Vibrio cholerae C7258, una cepa epidémica regional. Estos procedimientos permitieron la purificación y caracterización de preparaciones homogéneas de lactoferrina de alta pureza (98-99%), estructuralmente íntegra (92-93%), no tóxica, sin lipopolisacáridos (LPS) (menos de 0,4 EU/mg de proteína), con baja saturación de hierro (menos de 7% de saturación) y biológicamente activa (2,5 µM de lactoferrina inhibió el crecimiento in vitro de V. cholerae en 50%). La alta calidad de esta preparación de lactoferrina permite su empleo en estudios básicos de los mecanismos de acción bacteriana y en el desarrollo de nuevos medios para el diagnóstico de inflamaciones gastrointestinales, en los cuales se encuentra enfrascado nuestro grupo en estos momentos. Palabras claves: actividad antibacteriana, calostro humano, hierro, lactoferrina, LPS, purificación de proteínas, Vibrio cholerae Introduction Recognition of the immunoprotective potential of breast-feeding at epidemiological level [1], and the absence of similarly safe and effective pharmaceutical tools for prophylaxis of microbial infections in newborns [2] have promoted research on immunologically-active factors in human milk. Lactoferrin, a monomeric high-affinity non-haem iron-binding glycoprotein (calculated isoelectric point [pI] » 9.7, 77–80 kD), is present in the milk of all mammals except rabbits, rats and dogs [3]. In human milk, its concentration ranges from 37.5 to 87.5 µM in colostrum to about 12.5 µM in mature milk [4], from where it was firstly purified in 1960 [5–7]. It occurs in all body fluids at different concentrations, from higher values in secretions of exocrine glands [8] to lowest ones in human blood (~20 nM) [9, 10]. Its main biological role is to keep concentrations of free iron to amounts insufficient to support microbial growth in vivo, together with other high-affinity iron-binding glycoproteins. The significance of these proteins in host-parasite interactions and the mechanisms developed by microbial pathogens for iron uptake to overcome their effects, were recently reviewed [11, 12]. Lactoferrin has been involved in multiple effects in vitro, but no definitive corroboration of their physiological significance has yet been provided. To our knowledge, there are three structural properties directly related to its biological activity, which have also been used in purification strategies [9]. Firstly, lactoferrin comprises two homologous domains, each with one iron-binding site. Its antimicrobial effects are based on the capacity to withhold iron with high affinity even at low pH values (pH ~ 2.0), which can occur in certain body locations [12]. Its capacity to bind multivalent cations has been used for purification by metal-ion affinity chromatography [13]. Secondly, each lactoferrin domain comprises an N-linked complex biantennate glycosylation site whose main function seems to be proteolytic resistance [14]. Glycosylation differences allowed separation by concanavalin A affinity chromatography of bovine and human lactoferrins from the milk of transgenic cows [9]. Thirdly, non-specific binding of lactoferrin’s highly basic N-terminus to several eukaryotic and prokaryotic structures [15] has been implicated in antiviral effects against human immunodeficiency virus type 1 (HIV-1) [16], human cytomegalovirus (HCMV) [17], herpes simplex virus (HSV) types 1/2 [18], and hepatitis C virus (HCV) [19]. Immunomodulatory and anti-endotoxic effects mediated by direct binding to bacterial lipopolisaccharides (LPS) have been reported [20], as well as bactericidal effects due to interactions with anionic cell wall components of Gram-negative bacteria [21]. Cytokine-like activities ascribed to this protein have also been discussed [22]. The cationic nature of lactoferrin and the unique distribution of its superficial charges are widely used for purification by cation-exchange chromatography [9, 15]. Human milk lactoferrin has been used as a tool for research into the mechanisms of iron uptake by pathogens [12] and the potential of bacterial lactoferrin receptors for vaccine development has been mentioned [23]. The therapeutic potential of lactoferrin has been tested in animal models of diseases such as rheumatoid arthritis [24], staphylococcal kidney infection [25] and endotoxic shock [26], as well as in humans with chronic HCV infection [27]. Lactoferrin concentrations are a reliable biochemical marker for the diagnosis of bacterial infection-related diarrhoea [28], as well as gastrointestinal inflammation of autoimmune origin [29]. The realization by scientific, medical, and pharmaceutical communities of the therapeutic and prophylactic implications of iron metabolism in host-parasite relationships [30], along with the forthcoming availability of commercial recombinant lactoferrin [31, 32], suggest that there could be widespread clinical and research uses of this protein in the near future. In this context, the availability of a high-quality and homogenous lactoferrin preparation is crucial for the study of its multiple biological effects. Although several effective procedures for purification of lactoferrin from human milk have been developed [5–7, 9, 31], differences in biological activity —mainly due to the partial degradation of the N-terminus— and different degrees of iron saturation have been detected between commercial preparations. This indicates that there is a need for an accurate control of the structural properties directly related to the biological effects of lactoferrin [9, 31, 32]. On the other hand, the costs of commercially available lactoferrin preparations may preclude its use for research in some locations. Here, we describe practical procedures for the purification and characterization of non-toxic, highly-pure and functionally-active colostral lactoferrin, as well as techniques for the quality control of its most important structure-functional relationships. Materials and Methods Human colostrum Colostrum was kindly provided by healthy nursing women during the first five postpartum days at the Neonatological Units of Havana Gynecoobstetric Hospitals. All donors were seronegative to hepatitis B virus (HBV), HCV, HIV, Treponema pallidum and Toxoplasma gondii by the Diagnostic Services of the Mother-Infant Care Program of Cuba. Standard milk bank regulations were carefully followed to control microbial contamination [33]. Samples were stored at –20 ºC for no longer than three weeks before processing. Colostrum fractionation This was performed as previously reported [34] with major modifications. Frozen colostrum was thawed with running tap water at room temperature, diluted 9:1 (v:v) with 4.0 M NaCl, 0.1 M phosphate buffer (68.5 mM NaH2PO4, 31.5 mM Na2HPO4), adjusted to pH 4.6 with 0.1 M glacial acetic acid (Merck, Germany), and gently agitated for 1 h at 4 ºC. After centrifugation at 56,000 xg for 30 min at 4 ºC, three clearly separated layers were obtained: fats were in the top layer and casein plus cellular debris were at the bottom. The middle whey-containing layer was carefully collected and immediately adjusted to pH 6.5 with 0.1 M NaOH (Merck, Germany). Whey was further clarified by serial ultrafiltration using a glass fiber prefilter on top and three nitrocellulose membrane filters of 0.8, 0.45 and 0.2 µm pore-sizes placed on each other with polyester separators between them, as recommended by the manufacturer for difficult-to-filter liquids (Sartorious, Germany). The final whey was conveniently aliquoted in glass containers and stored at –20 ºC until use. Chromatographic processing A XK-50 column (Pharmacia-LKB, Sweden) packed with 100 mL of a strong cationic exchanger (SP Sepharose Fast Flow, Pharmacia-LKB, Sweden) and equilibrated with 0.4 M NaCl, 0.01 M phosphate buffer, pH 6.50, was used for cation-exchange chromatography. Lactoferrin purification was performed essentially as reported [9, 29] with some variations. Clarified whey was passed through the column at a linear flow rate of 0.67 cm/min. Bound lactoferrin was eluted in a single peak with 0.8 M NaCl, 0.01 M phosphate buffer, pH 6.5. This fraction was concentrated against 0.01 M phosphate buffer, pH 6.50, and desalted to 0.15 M NaCl in an Amicon ultrafiltration stirred cell with controlled nitrogen supply (Amicon, USA). Lactoferrin samples were aliquoted and stored either frozen at –20 ºC or at 4 ºC after conventional freeze-drying (Cryodos Telstar, Spain). Eluates were saved and used as source for purification of secretory IgA (Center for Immunoassays, Cuba, data not shown). Final ultrafiltration, filling and sealing steps were carried out under aseptic conditions. The Laboratory for Analytical Technical Control (Finlay Institute, Cuba) tested colostral whey and lactoferrin samples for microbial sterility. Analytical methods Protein concentrations were determined by the method of Lowry [35] using bovine serum albumin (BSA) as standard (BDH, UK). Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was routinely performed in a discontinuous gel system [36]. All samples were boiled for 5 min under reducing conditions and 60 µg of total proteins were applied onto 12.5% polyacrylamide gels. Human lactoferrin and molecular weight markers (Sigma, USA) were used for antigenic identity and molecular weight estimation, respectively. Quantification of relative concentrations and molecular weight estimation of the bands antigenically identified as lactoferrin were performed by densitometry of silver nitrate (Sigma, USA)-stained gels [37] using an in-built software (Laser UltraScan XL Densitometer, Pharmacia-LKB, Sweden). To ensure that all bands were taken into account, the low density rejection function was set to zero. Western blotting was performed as reported elsewhere [38]. After electrophoresis, gels were soaked in TBS (Tris-buffered saline: 50 mM Tris-HCl, 150 mM NaCl, pH 8.0) to reduce SDS concentration and incubated in transfer buffer (20 mM Tris, 150 mM glycine, 20% methanol, 0.1% SDS, pH 8.3) for 10 min. After transfer for 2 h at 4 ºC, 0.2-µm nitrocellulose membranes (Pharmacia-LKB, Sweden) were incubated in TBS-T blocking solution (TBS plus 0.05% Tween 20, pH 8.0) containing 1% BSA Fraction V (Sigma, USA). After three washes, membranes were incubated in primary binding solution containing 1:1000 TBS-T-diluted anti-human lactoferrin rabbit polyclonal antibodies. After three washings, the membranes were incubated in a secondary binding solution containing horseradish peroxidase (HRPO)-conjugated goat anti-rabbit IgG (SAPU, Scotland) diluted 1:2000 in TBS-T. This mixture was removed, the membranes were washed five times, and then exposed to HRPO substrate solution (0.02% [w/v] 4-chloro-1-naphtol [Sigma, USA] and 0.06% [v/v] hydrogen peroxide [BDH, UK] in TBS) until color developed. The reaction was stopped by washing out the substrate solution with distilled water. To check for possible contamination with IgA [39], Western blotting was performed as described above, but after the blocking step, nitrocellulose membranes were directly exposed to HRPO-conjugated goat anti-human IgA (SAPU, Scotland) diluted 1:2000 in TBS. Human IgA (Sigma, USA) was used as positive control (data not shown). Incubation steps were performed for 1 h at 37 ºC with gentle agitation. N-terminus integrity of purified lactoferrin Lactoferrin was characterized by the pattern of elution in cation-exchange high-performance liquid chromatography (HPLC) [14]. Freeze-dried lactoferrin was diluted to 500 µg/mL in 20 mM Tris pH 7.5 and applied onto a Mono S HR 5/5 HPLC column (Pharmacia, Sweden) at a linear flow rate of 5.0 cm/min. The protein was eluted with a linear salt gradient of 0.0–1.0 M NaCl. Peaks detected at 226 nm were collected and characterized by SDS-PAGE and Western blotting as described above. Chromatographic parameters were determined with Biocrom programme, version 2.3 (Automation Division, Center for Genetic Engineering and Biotechnology, Cuba). As reference, commercial lactoferrin (Sigma, USA) was assayed under the same conditions. Endotoxin contamination Protein samples were tested by the Limulus amebocyte lysate assay (Chromogenic CoatecÒ AB, Sweden). A value of 0.25 EU/mg of protein was the limit of acceptance established. Preparation of lactoferrin derivatives Apolactoferrin, the iron-free form, was produced as reported [40]. Briefly, the protein was diluted to 1% (w/v) in 0.1 M sodium phosphate, 0.1 M sodium acetate, 40 mM EDTA, and adjusted to pH 4.0 by dropwise addition of concentrated glacial acetic acid. The solution was firstly equilibrated and dialyzed against the same buffer overnight at 4 ºC. Hololactoferrin, the 100% iron-saturated form, was obtained by incubation of 1% lactoferrin (w/v) in a ferric nitrilotriacetic acid (FeNTA) solution (Fe:NTA molar ratio 1:4) overnight at 20 ºC. To ensure full iron saturation, a twofold molar excess of FeNTA per iron binding site was added to lactoferrin solution. In both procedures, final samples were extensively dialyzed against 0.15 M NaCl, 0.01 M phosphate buffer, pH 6.5. Iron saturation was judged by comparing absorbance values at 280 and 465 nm [14, 40]. Preparation of lactoferrin antiserum Male New Zealand adult white rabbits were subcutaneously immunized with 0.05 mg of human lactoferrin (Sigma, USA) using Freund’s complete adjuvant (Difco Laboratories, USA) in the first injection. The same dose and route were used at 2, 4, and 6 weeks but in Freund’s incomplete adjuvant (Difco Laboratories, USA). Terminal bleeds were performed 2 weeks after the final injection and the antiserum was found to be adequate for the specific identification of lactoferrin by Western blotting at a 1:2000 dilution. The antiserum was aliquoted and stored at –20 ºC. Bacterial culture medium Chemicals of the highest purity were purchased from Sigma, USA. Chemically-defined medium (CDM) was prepared as previously reported [41] with some modifications [42]: 59 mM glucose, 12 mM Na2HPO4, 69 mM NaCl, 9.3 mM NH4Cl, 7.3 mM KH2PO4, and 2.8 mM MgSO4. Aminoacids were added to the following final concentrations (mg/L): glutamic acid, 1200; aspartic acid, 1200; tyrosine, 100; tryptophan, 30; cystein, 120; isoleucine, 300; valine, 240; methionine, 90; phenylalanine, 100; leucine, 300; glycine, 1200; alanine, 1200; proline, 1200; lysine, 300; histidine, 240; arginine, 360; threonine, 2400; and serine, 2400. Vitamins were added to the following final concentrations (mg/L): nicotinic acid, 0.5; calcium-d-pantothenate, 0.5; thiamine, 0.5; biotin, 0.003. A mineral micronutrients solution was also added to the following final concentrations (µM): CaCl2·6H2O, 0.5; CoCl2·6H2O, 0.05; CuSO4·5H2O, 0.05; ZnSO4·7H2O, 0.05; MnSO4, 0.1; (NH)Mo7O24·4H2O, 0.005. For iron deprivation, all components except vitamins and MgSO4 were deferrated by stirring for 3 h with 20% iminodiacetic acid (Chelex 100 chelating resin, Sigma, USA). Final pH was adjusted between 7.0 and 7.1. Sterile disposable plasticware (Corning, USA) was always used to prevent contamination with iron. Bacterial strain and growth conditions Vibrio cholerae wild type strain C7258 El Tor, Ogawa (1991 Peruvian outbreak isolate, RA Finkelstein, University of Missouri, USA), stored at –70 ºC in skim milk (Oxoid Ltd, UK) and supplemented with 15% glycerol, was streaked onto thiosulfate citrate bile salts (Oxoid Ltd, UK) agar plates and grown aerobically for 12 h at 37 ºC. To reduce stored iron levels, a colony was inoculated in 1 mL of CDM and pre-cultured overnight in a 5-mL plastic tube at 250 rpm and 37 ºC in a shaker incubator (G76D New Brunswick Scientific, USA). The culture was harvested (6000 xg, 10 min, 4 ºC), washed with CDM and used to inoculate fresh CDM cultures to 0.1 optical density (OD) unit at 600 nm. Growth was monitored by determining the absorbance at 600 nm of samples taken at 60-min intervals. The same culture conditions were used in all the experiments. Lactoferrin antibacterial activity As we previously reported for Staphylococcus aureus [42], the minimal concentration of lactoferrin capable of inhibiting the growth of V. cholerae to 50% during exponential phase (MIC50%) was determined by the addition to CDM of lactoferrin concentrations between 0.3 µM and 20 µM at the time of inoculation. Lactoferrin-free CDM was used as negative control. To confirm the bacteriostatic effect of iron-deprivation upon the growth of V. cholerae C7258 strain, an identical culture was supplemented with 800 µM EDDHA (Sigma, USA), a specific ferrochelating reagent. Cultures were observed for 10 h. Iron reversibility of lactoferrin antibacterial effect The iron reversibility of the growth inhibitory activity of lactoferrin against V. cholerae was tested by adding a 20-fold molar excess of FeNTA at 0, 3 and 6 h, respectively, to three different CDM cultures which were supplemented with MIC50% lactoferrin at the time of inoculation. As control, non-iron supplemented but lactoferrin-containing CDM cultures were kept under similar conditions. Statistical analysis All experiments described were performed at least three times, and the standard error, the mean and the coefficient of variation were calculated for all data. The mean values for OD measurements in all cultures were compared with the control culture and among them by the Student’s t test. Values of P < 0.05 were considered statistically significant. Results Human colostrum All colostrum samples retained their typical organoleptic properties. A pH value of 7.0 ± 0.2 was consistently measured in these samples, which positively correlated with the absence of microbial contamination as observed in the microbial sterility assays (data not shown). Colostrum fractionation In a previous work, we observed that colostral lactoferrin bound to an SP Sepharose FF matrix consistently eluted as a single peak at 0.6–0.7 M NaCl in a 0.0 M–1.0 M NaCl linear gradient. No more peaks were detected thereafter, even at ionic strengths of up to 2.0 M NaCl [33]. As a result, ionic strength and phosphate concentration in whey samples were increased to 0.4 M NaCl and 0.01 M phosphate, respectively, before acid precipitation, and the cationic matrix was equilibrated with the same buffer composition. Addition of 0.4 M NaCl and 0.01 M phosphate to colostrum samples practically eliminated precipitation of lactoferrin in the casein fraction. Lactoferrin concentration in whey fractions rose by 20–30%, compared to non-treated samples from the same batch. Chromatographic processing Previous treatment with 0.4 M NaCl increased the efficiency of the chromatographic process by allowing nearly the triplication of the volume of whey to be applied per volume of bed—1.0 L vs. 0.35 L of whey per 100 mL of resin—without breakthrough occurrence [33] and reducing the time for chromatographic processing in about 50%. Lactoferrin consistently eluted at 0.8 M NaCl in a single symmetric peak at a 1.5 ± 0.1 bed volume (Figure 1). The protein yield represented 83–85% of the initial concentration, nearly the double of the values reached when lactoferrin was directly isolated from the whey samples (3.6 g vs. 6.0 g per liter of colostrum) [33]. Endotoxin levels were significantly lower in the samples isolated from treated vs. non-treated colostral whey, and were within the limit of acceptability—less than 0.4 EU/mg of protein. Taken together, these results indicate a complete disruption of the non-specific interactions between lactoferrin and caseins during acid precipitation, as well as a separation from LPS and other contaminants due to its higher affinity for SP Sepharose resin at 0.4 M NaCl. Figure 1. Pattern of elution of lactoferrin purified from human colostral whey from SP Sepharose FF Matrix. For specific conditions see the text. Densitometric analysis of electrophoretically separated protein bands consistently showed a major band at 78–80 kD, which represented 98–99 % of the total protein applied per lane. Few lower molecular weight bands were detected in both eluted and Sigma lactoferrin samples (Figure 2). These bands were antigenically identified as degraded forms of lactoferrin [4, 9]. Figure 2. Patterns of processed human colostral whey (lane 1), eluate (lane 2), eluted lactoferrin (lane 3), and Sigma lactoferrin (lane 4) after electrophoretic separation and silver staining in a 12.5% SDS-PAGE. Observe the similarities in the electrophoretic profiles of the two lactoferrin samples. Estimated molecular weights (kD) are shown at the left side. Contrary to a previous report [39], no IgA contamination could be detected, even in very concentrated lactoferrin samples purified from treated colostral whey, while minor non-quantified IgA contamination was detected when non-treated colostral whey was used [33, 34] (data not shown). Mono S HR HPLC separation of lactoferrin samples indicated that 92.8 ± 1.12% of the protein eluted between 0.6 M and 0.69 M NaCl. Three minor peaks eluted at 0.17 M, 0.5 M and 0.56 M NaCl, which altogether represented less than 7% of the total protein eluted. No statistically significant differences in the levels of purity and the pattern of elution from Mono S HR HPLC were detected between the commercial reference and the preparation assayed under similar conditions. Lactoferrin from Sigma and the samples eluted at 0.615 M and 0.624 M NaCl, and at 64.04 and 64.88-min retention times, respectively (Figure 3). Apo and iron-saturated forms of purified lactoferrin were obtained as described above, and the ratio of absorbance values at 465 and 280 nm of a 1% lactoferrin solution was in agreement with previously reported values; i.e. 0.60 for 100% iron-saturated lactoferrin and nearly zero for apolactoferrin [40]. No differences in absorbance were found for the lactoferrin from Sigma, either in the apo or in the holo forms (data not shown). Figure 3. Chromatograms of commercial (upper) and colostrum-purified (lower) lactoferrin samples analysed by Mono S HPLC under identical conditions. No differences in retention time (64.04 min vs. 64.88 min) and molarity of elution (0.615 M vs. 0.624 M) between samples could be detected. The solid diagonal line represents the linear saline elution gradient from 0.0 M to 1.0 M NaCl. Antibacterial activity The growth of V. cholerae strain C7258 in CDM was inhibited by the addition of lactoferrin in a dose-dependent manner. The inhibitory effect was either undetectable or only transiently observed at concentrations below 1.25 µM. The minimal concentration of lactoferrin which consistently reduced V. cholerae growth in 50% during the logarithmic phase was 2.5 µM. This effect was saturable as no significantly higher growth inhibition could be achieved by increasing lactoferrin concentration up to 8 times above this value (Figure 4). A greater and more stable inhibition was observed in cultures treated with 800 µM EDDHA, which corroborated the requirement of iron for the normal growth of this strain under the conditions of this study [12, 43]. Figure 4. Concentration dependence of lactoferrin microbiostatic effect against Vibrio cholerae C7258. A 2.5-µM lactoferrin consistently halved bacterial growth during logarithmic phase. The solid horizontal line represents 50% of growth inhibition. This effect was saturable, as no further growth inhibitory effect was observed for lactoferrin concentrations up to eight times higher (20 µM). Iron reversibility of lactoferrin antibacterial effect A 20-fold molar excess of FeNTA, i.e. 50 µM, completely reversed the growth inhibitory effect of MIC50% lactoferrin (2.5 µM). Interestingly, it was noted that the kinetics of iron reversion seemed to be affected by the time of exposure of the bacterial cultures to lactoferrin. A longer delay before resumption of normal growth after addition of FeNTA was consistently observed in cultures treated with 2.5 µM lactoferrin for more than 4–5 h. This effect does not seem to be dose-dependent, as a similar delay was observed at higher lactoferrin concentrations (data not shown). We did not observe this effect in previous experiments using S. aureus strains [42]. It suggests possible differences in the microbiostatic activity of lactoferrin against V. cholerae. Discussion Since lactoferrin is the most cationic protein present in human milk, its non-specific electrostatic interactions with anionic moieties of molecules such as caseins, lysozyme, IgA secretory component, a-lactoalbumin, and b-lactoglobulin [9, 14, 33], as well as bacterial LPS [20], are disrupted only at ionic strengths above 0.3 M NaCl. Consequently, addition of up to 0.4 M NaCl and 0.01 M phosphate prior to the delipidation and casein precipitation steps prevents coprecipitation with caseins and increases the yield and purity of lactoferrin before chromatographic processing. Because lactoferrin has the highest affinity for cationic exchange resins and a clustered distribution of its positive charges at the N-terminus, it can be selectively separated in a substantially pure, intact and complex-free form in a single chromatographic step at a high ionic strength. [14, 33]. Therefore, lactoferrin preparations were consistently 98–99% pure and essentially free of LPS contamination (below 0.4 EU/mg of protein). The structural integrity of the purified protein is not affected (Figure 3), as shown by the pattern of elution in Mono S HPLC, a reliable and more affordable technique for monitoring the degree of structural integrity of lactoferrin preparations as previously reported [9]. The lack of toxicity of this preparation was confirmed in two different experimental settings. Doses as high as 30 mg were injected intravenously into adult Balb/c mice without causing any detectable clinical or histopathological damage (data not shown). Concentrations as high as 4–5 mg/mL of lactoferrin were required before any cytotoxicity against cultured Vero cells was observed (data not shown). Lactoferrin concentrations 30-fold lower than those present in human colostrum showed a significant bacteriostatic activity against a regional epidemic strain of V. cholerae. This demonstrates the biological activity of these preparations, corroborates the potential of lactoferrin as a broad-spectrum natural antimicrobial agent, as shown both in vitro [16–21, 31] and in vivo [24–27], and reinforces the importance of breast-feeding for the prevention of gastrointestinal microbial infections in newborns [1]. As with other El Tor strains, V. cholerae C7258 strain secretes the hemagglutinin/metalloenzyme Hap with proteolytic activity against lactoferrin and other protective components of the immune system [44]. Clinical trials using genetically-attenuated V. cholerae CTXf- vaccine strains seem to implicate Hap in cholera virulence, as detection of intestinal inflammatory effects and increased levels of lactoferrin and proinflammatory cytokines were limited to stools of human volunteers immunized with a hap+ strain [45], but not with the parental hap- strain ([46], García L, Finlay Institute, Havana, personal communication). In contrast, Toma et al. reported that the proteolytic degradation of lactoferrin by Hap does not affect its antimicrobial activity in vitro against a non-O1 strain of V. cholerae isolated from humans [47]. We observed that V. cholerae, but not the S. aureus strains assayed under similar conditions, showed a direct correlation between the time of exposure to lactoferrin and the delay in resumption of normal growth after addition of iron [42]. Experiments are being carried out to elucidate the actual effect of Hap-mediated proteolysis on the mechanisms of antimicrobial activity of lactoferrin against V. cholerae, and on the mechanisms of iron uptake by V. cholerae strains in vivo. We conclude that the procedures described in this article for the purification and quality control of lactoferrin provide fast, simple and state-of-the-art methods for obtaining a high quality, substantially pure, structurally intact, and biologically functional lactoferrin from human colostrum. This lactoferrin preparation shows quality control parameters equivalent to commercially available preparations. Therefore, this lactoferrin preparation may be used in basic studies on the mechanisms of microbial virulence related to iron metabolism and in the development of new tools for the diagnosis of gastrointestinal inflammation of infectious or autoimmune origin. Acknowledgments We thank Vivian Lago, Odalys Guerra and Alejandro Ramírez, from the Development Unit of the Finlay Institute for their technical assistance in colostrum processing and freeze-drying, respectively; Dayris Hernández and Marlenis Cardoso, from the Vaccine Manufacturing Plant, Finlay Institute, for endotoxin determinations. We are deeply indebted to Damián Castellanos, Faculty of Biology, University of Havana, for his contribution to the experimental work; and to physicians and nurses from the Neonatological Services of Havana Ginecoobstetric Hospitals América Arias, Clodomira Acosta, and Ramón González Coro, for their collaboration in colostrum collection and kind encouragement. Collaboration between JHB and AAL is supported by grants from The Royal Society, United Kingdom. References 1. Hanson LÅ, Telemo E. Immunobiology and epidemiology of breastfeeding in relation to prevention of infections from a global perspective. In: Ogra PL, Mestecky J, Lamm ME, Strober W, Bienenstock J, McGhee JR, editors. Mucosal immunology. 2nd ed. San Diego: Academic Press; 1999. p.1501–10. 2. Kinney J, Mundorf L, Gleason C, Lee C, Townsend T, Thibault R, et al. Efficacy and pharmacokinetics of intravenous immune globulin administration to high-risk neonates. Am J Dis Children 1991;145:1233–8. 3. Masson PL, Heremans JF. Lactoferrin in milk from different species. Comp Biochem Physiol 1971;39B:119–29. 4. Goldman AS, Garza C, Nichols BL, Goldblum RM. Immunologic factors in human milk during the first year of lactation. J Pediatrics 1982;100:563–7. 5. Montreuil J, Tonnelat J, Mullet S. [Preparation and properties of lactosiderophilin (lactotransferrin) from human milk. French]. Biochim Biophys Acta 1960; 45:413–21. 6. Groves ML. The isolation of a red protein from milk. J Amer Chem Soc 1960;82: 3345–50. 7. Johanson B. Isolation of an iron-containing red protein from human milk. Acta Chem Scand 1960;14:510–3. 8. Masson PL, Heremans JF, Dive Ch. An iron-binding protein common to many external secretions. Clin Chim Acta 1966; 14:735–9. 9. Nuijens JH, van Berkel PHC, Schanbacher FL. Structure and biological actions of lactoferrin. J Mam Gland Biol Neopl 1995;1:285–95. 10. McAbee DD, Esbensen K. Binding and endocytosis of apo- and holo-lactoferrin by isolated rat hepatocytes. J Biol Chem 1991;266:23624–31. 11. Bullen JJ, Griffiths E. Iron-binding proteins and host defences. In: Bullen JJ, Griffiths E, editors. Iron and infection. New York: John Wiley & Sons Ltd; 1999. p.327–450. 12. Griffiths E, Williams P. The iron-uptake systems of pathogenic bacteria, fungi and protozoa. In: Bullen JJ, Griffiths E, editors. Iron and infection. New York: John Wiley & Sons Ltd; 1999. p.87–212. 13. Kågedal L. Immobilized metal ion affinity chromatography. In: Janson J-Ch, Rydén L, editors. Protein purification: principles, high-resolution methods, and applications. 2nd ed. New York: John Wiley & Sons, Inc. 1998. p.311–42. 14. van Berkel PHC, Geerts MEJ, van Veen HA, Kooiman PM, Pieper FR, de Boer HA, Nuijens JH. Glycosylated and unglycosylated human lactoferrin both bind iron and show identical affinities towards human lysozyme and bacterial lipopolysaccharide, but differ in their susceptibility towards tryptic proteolysis. Biochem J 1995;312:107–14. 15. van Berkel PHC, Geerts MEJ, van Veen HA, Mericskay M, de Boer HA, Nuijens JH. N-terminal stretch Arg2, Arg3, Arg4, and Arg5 of human lactoferrin is essential for binding to heparin, bacterial lipopolysaccharide, human lysozyme and DNA. Biochem J 1995;328:145–51. 16. Viani RM, Gutteberg TJ, Lathey JL, Spector SA. Lactoferrin inhibits HIV-1 replication in vitro and exhibits synergy when combined with zidovudine. AIDS 1999; 13:1273-1274. 17. Harmsen MC, Swart PJ, de Béthune M-P, Pauwels R, De Clercq E, The TH, et al. Antiviral effects of plasma and milk proteins: lactoferrin shows potent activity against both human immunodeficiency virus and human cytomegalovirus replication in vitro. J Infect Dis 1995;172: 380–8. 18. Marchetti M, Pisani S, Antonini G, Valenti P, Orsi N. Metal complexes of bovine lactoferrin inhibit the in vitro replication of herpes simplex virus type 1 and 2. Biometals 1998;11/12:89–94. 19. Ikeda M, Nozaki A, Sugiyama K, Tanaka T, Naganuma A, Tanaka K, et al. Characterization of antiviral activity of lactoferrin against hepatitis C virus infection in human cultured cells. Vir Res 2000;66:51–63. 20. Elass-Rochad E, Legrand D, Salmon V, Roseanu A, Trif M, Tobias PS, et al. Lactoferrin inhibits the endotoxin interaction with CD14 by competition with the lipopolysaccharide-binding protein. Infect Immun 1998;66:486–91. 21. Ellison RT III. The effects of lactoferrin on Gram-negative bacteria. In: Hutchens TW, Rumball SV, Lönnerdal B, editors. Lactoferrin: structure and function. New York: Plenum Press; 1994. p.71–90. 22. Brock JH. Lactoferrin. In: Aggarwal BB, editor. Human cytokines. Handbook for basic and clinical research. Maryland: Blackwell Science; 1999. p.92–123. 23. Schryvers AB, Bonnah R, Yu R, Wong H, Retzer M. Bacterial lactoferrin receptors. In: Spik G, et al, editors. Advances in lactoferrin research. New York: Plenum Press; 1998. p.123–33. 24. Guillen C, McInnes IB, van Verkel PHC, Brock JH. Anti-inflammatory effect of lactoferrin in mouse models of arthritic disease. In: Shimazaki K, Tsuda H, Tomita M, Kuwata T, Perraudin JP, editors. Lactoferrin: structure, function and applications. Amsterdam. Elsevier Science BV; 2000. p.103–9. 25. Bhimani RS, Vendrov Y, Furmanski P. Influence of lactoferrin feeding and injection against systemic staphylococcal infection in mice. J App Microb 1999;86:135–44. 26. Lee WJ, Farmer JL, Hilty M, Kim YB. The protective effects of lactoferrin feeding against endotoxin lethal shock in germfree piglets. Infect Immun 1998;66:1421–6. 27. Tanaka K, Ikeda M, Nozaki A, Kato N, Tsuda H, Saito S, Sekihara H. Lactoferrin inhibits hepatitis C virus viremia in patients with chronic hepatitis C. In: Shimazaki K, Tsuda H, Tomita M, Kuwata T, Perraudin JP, editors. Lactoferrin: structure, function and applications. Amsterdam. Elsevier Science BV; 2000. p.361–7. 28. Choi SW, Park CH, Silva TMJ, Zaenker EI, Guerrant RL. To culture or not to culture: fecal lactoferrin in screening for inflammatory bacterial diarrhea. J Clin Microbiol 1996;34:928–32. 29. Fine KD, Ogunji F, George J, Niehaus MD, Guerrant RL. Utility of a rapid fecal latex agglutination test detecting the neutrophil protein, lactoferrin, for diagnosing inflammatory causes of chronic diarrhea. Am J Gastroenterol 1998;93:1300–5. 30. Griffiths E, Bullen JJ. Iron and Infection: future prospects. In: Bullen JJ, Griffiths E, editors. Iron and infection. New York: John Wiley & Sons Ltd; 1999. p.451–81. 31. Ward PP, Piddington GA, Cunningham GA, Zhou X, Wyatt RD. A system for the production of commercial quantities of human lactoferrin: a broad spectrum natural antibiotic. Bio/Technology 1995; 13:498–503. 32. Headon DR. Human lactoferrin: production at large scale, characterization and applications. In: Shimazaki K, Tsuda H, Tomita M, Kuwata T, Perraudin JP, editors. Lactoferrin: structure, function and applications. Amsterdam. Elsevier Science BV; 2000. p.415–27. 33. Castellanos García D. Purificación y caracterización de la lactoferrina humana. [Tesis]. Habana: Facultad de Biología. Universidad de La Habana; 1999. 34. Tomasi TB, Tan EM, Solomon A, Prendergast A. Characteristics of an immune system common to certain external secretions. J Exp Med 1965;121:101–24. 35. Lowry OH, Rosenbourgh NJ, Farr AL, Randall RJ. Protein measurement with the Folin phenol reagent. J Biol Chem 1951; 193:265–75. 36. Laemmli UK. Cleavage of structural protein during assembly of the head of bacteriophage T4. Nature 1970;277: 680–5. 37. Sambrook J, Fritsch EF, Maniatis T. Molecular cloning. A laboratory manual. 2nd ed. New York: Cold Spring Harbor Laboratory Press; 1989. p.18–56. 38. Towbin H, Staehelin T, Golden J. Electrophoretic transfer of proteins from polyacrylamide gel to nitrocellulose sheets procedure and some applications. Proc Natl Acad Sci 1979;76:4350–4. 39. Watanabe T, Nagura H, Watanabe K, Brown WR. The binding of human milk lactoferrin to immunoglobulin IgA. FEBS Letters 1984;168:203–7. 40. Mazurier J, Spik G. Comparative study of the iron-binding properties of human transferrins. Bioch Bioph Acta 1980;629: 399–408. 41. Kadurugamuwa JL, Anwar H, Brown MRW, Shand GH, Ward KH. Media for study of growth kinetics and envelope properties of iron-deprived bacteria. J Clin Microbiol 1987;25:849–55. 42. Aguila A, Guillén C, de Lillo A, Brock JH. Inhibition of growth of Staphylococcus aureus is a function of its iron-binding properties. In: Gavilondo JV, Ayala M, Acevedo B, editors. Biotecnología Habana´99. Avances en Biotecnología Moderna; 1999 Nov 28-Dec 3; La Habana, Cuba. La Habana: Elfos Scientiae; 1999. p.N9. 43. Tashima KT, Carroll PA, Rogers MB, Calderwood SB. Relative importance of three iron-regulated outer membrane proteins for in vivo growth of Vibrio cholerae. Infect Immun 1996;64:1756–61. 44. Finkelstein RA, Boesman-Finkelstein M, Holt P. Vibrio cholerae hemagglutinin/lectin/protease hydrolyzes fibonectin and ovomucin: F.M. Burnet revisited. Proc Natl Acad Sci USA 1983;80:1092–5. 45. Silva TMJ, Schleupner MA, Tacket CO, Steiner TS, Kaper JB, Edelman R, Guerrant RL. New evidence for an inflammatory component in diarrhea by selected new, live attenuated cholera vaccines and by El Tor and O139 Vibrio cholerae. Infect Immun 1996;64:2362–4. 46. Benítez JA, García L, Silva A, García H, Fando R, Cedré B, et al. Preliminary assessment of the safety and immunogenecity of a new CTXf-negative, hemagglutinin/protease-defective El Tor strain as a cholera vaccine candidate. Infect Immun 1999; 67:539–45. 47. Toma C, Honma Y, Iwanaga M. Effect of Vibrio cholerae non-O1 protease on lysozyme, lactoferrin and secretory immunoglobulin A. FEMS Microbiol Lett 1996; 135:143–7. Copyright Elfos Scientiae 2000 The following images related to this document are available:Photo images[ba00052c.jpg] [ba00052b.jpg] [ba00052a.jpg] [ba00052d.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}