 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnología Aplicada 2000;17:215-220
Respuesta inmunológica contra garrapatas
Manuel Rodríguez Valle
División de Genética de Células de Mamíferos. Centro de Ingeniería
Genética y Biotecnología.
Recibido en noviembre de 1999. Aprobado en abril del 2000.
Code Number: ba00068
RESUMEN
Las interacciones hospedero-parásito se caracterizan por ser muy complejas. Las garrapatas inducen una respuesta inmunológica reguladora en el hospedero y reacciones efectoras que involucran anticuerpos, el sistema del complemento, células presentadoras de antígenos, linfocitos T y otras biomoléculas activas. Las garrapatas contrarrestan las defensas del hospedero mediante la reducción de la proliferación de linfocitos T; de la producción de IL-2, IFN-g, IL-1, TNF; y de la respuesta de anticuerpos. El balance dinámico entre la resistencia adquirida por el hospedero y la modulación de dicha resistencia por los componentes salivales de la garrapata, puede inducir cambios que afectan el desarrollo del ectoparásito y su capacidad de transmisión de patógenos. Un considerable esfuerzo se realiza para desarrollar vacunas que induzcan inmunidad en los animales contra los ectoparásitos. La identificación de antígenos esenciales y su producción como proteínas recombinantes eficientes aún representa un obstáculo. En este trabajo, se discuten las características de algunos antígenos y el modo de acción de la respuesta inmune inducida por ellos para controlar las poblaciones de garrapatas. El desarrollo de nuevas vacunas contra las garrapatas depende del reconocimiento como moléculas blanco, de antígenos similares a los descritos en este trabajo. Por lo tanto, la eficacia de estas vacunas dependerá no sólo de las características del antígeno, sino también de su función en la biología del sistema digestivo o en otros órganos de la garrapata. A partir de la comprensión de la inmunidad adquirida y de las respuestas inducidas por los antígenos ocultos contra las garrapatas, será posible desarrollar racionalmente una vacuna efectiva contra las garrapatas.
Palabras claves: Boophilus microplus, garrapata, inmunomodulación, vacuna
ABSTRACT
Immune Response to Cattle Tick. Tick-host relationship is characterized by complex immunological interactions. Tick feeding induces host immune regulatory pathways involving antibodies, complement, antigen-presenting cells, T lymphocytes, and other bioactive molecules. Tick countermeasures to host defenses reduce T-lymphocyte proliferation; production of IL-2, IFN-g, IL-1 and tumor necrosis factor (TNF); and antibody responses. The dynamic balance between acquired resistance and tick modulation of host immunity affects engorgement and pathogen transmission. Considerable efforts have led to the development of a vaccine to induce animal immunity to the parasite. The identification of critical antigens and their production as effective recombinant proteins remain the greatest hurdle. The characteristics of the few known antigens and the mode of action of the protective immune response are discussed. Development of further vaccines will depend on the recognition of likely antigenic targets. The efficacy of such vaccines will depend on the characteristics of the target species, in particular its digestional biology. Thorough understanding of the acquired and induced immunity to tick will make possible the rational development of anti-tick vaccines.
Keywords: Boophilus microplus, immunomodulation, tick, vaccine
Introducción
El bovino es el hospedero principal del ectoparásito Boophilus microplus, el cual es vector de enfermedades que atacan al ganado vacuno y está distribuido geográficamente por todas las regiones tropicales y subtropicales del planeta, por lo que se ha convertido en un grave problema para la ganadería. Las mermas principales que sufren los criadores de ganado bovino por la incidencia de este ectoparásito, son la muerte de los animales y la disminución de la producción de carne y leche. Por ejemplo, en 1982, fallecieron en Brasil 2,5 millones de cabezas de ganado, lo cual representó la pérdida de 75 millones de kilogramos de carne, 1,5 billones de litros de leche, 8,6 millones de dólares por daños secundarios, y 25 millones de dólares en acaricidas químicos para combatir las infestaciones por garrapatas [1]. En Australia, las pérdidas en la producción ganadera a causa de las garrapatas se estiman entre cien y cientocincuenta millones de dólares al año [2], y en Sudamérica se calcula que están cerca de mil millones de dólares anuales [1]. En 1985, las pérdidas en Cuba por las garrapatas ascendían a 25 millones de pesos y durante 1986 existía un consumo de unos cinco millones de dólares anuales en acaricidas químicos, sin que esta suma llegara a satisfacer las necesidades.
El método rutinario para controlar este ácaro es el tratamiento del ganado vacuno con acaricidas químicos. Sin embargo, este método de lucha presenta numerosas desventajas en la actualidad, entre las cuales la más importante es la creciente aparición de poblaciones de ácaros resistentes al efecto tóxico de las sustancias químicas. Los mejores ejemplos se pueden encontrar en Australia, donde existen garrapatas que son resistentes a sustancias como el arsénico, el DDT, compuestos organoclorados, una gama de compuestos organofosforados, organocarbamatos, amidinas y, finalmente, piretroides sintéticos [3]. En México, se han aislado cepas doble y triple resistentes a compuestos organofosforados, clorados y piretroides, como por ejemplo, las cepas denominadas Tuxpan y Mora. Todo esto multiplica las dificultades de lucha contra las garrapatas y muestra un futuro poco alentador para el uso de productos químicos en la erradicación de los ectoparásitos.
Otro elemento que puede influir en el uso de los acaricidas químicos es la falta de recursos para continuar los tratamientos, sobre todo en los países en vías de desarrollo, situación que deja a los animales extremadamente susceptibles a las garrapatas y a las enfermedades que ellas transmiten. Por otra parte, en el mundo de hoy el hombre se ha percatado del grave problema que constituye consumir alimentos que contengan residuos químicos. Esto ha provocado que se destinen millones de dólares anualmente a supervisar la presencia de residuos químicos en las carnes y la leche, con el objetivo de acatar las estrictas regulaciones internacionales, tanto para las exportaciones como para el consumo nacional de estos alimentos. Por lo tanto, queda bien clara la urgente necesidad de encontrar nuevas opciones de lucha para el control de los ectoparásitos en general, que permitan un uso racional de los acaricidas químicos. Estas nuevas formas de lucha deberán eliminar el carácter tóxico de las formas tradicionales de lucha para, de cierta forma, dejar de afectar la salud de los animales, el hombre y el medio ambiente.
La vacunación contra las garrapatas como método de control
Los problemas que han presentado los productos químicos durante las últimas décadas, han motivado que se centre la atención científica en la búsqueda de nuevos métodos para el control de los ectoparásitos. El método inmunológico es capaz de inducir una respuesta tal en los animales vacunados, que permite mantener controlados a estos parásitos. Esta vía tiene como perspectiva una protección de mayor duración y está exenta de problemas de índole ambiental, a diferencia del tratamiento con acaricidas químicos. Los antecedentes de este método se manifiestan en la naturaleza, donde existen casos de control inmunológico. Después de una infestación natural reiterada por el parásito, algunos animales presentan una resistencia adquirida que provoca una disminución sustancial del número de garrapatas por animal. Este resultado es muy utilizado en la explotación ganadera actual, aunque se sabe que el grado de resistencia adquirida varía mucho entre las razas bovinas. Un ejemplo lo constituye el cruzamiento entre Bos indicus (Cebú) y Bos taurus (Holstein) para generar un F1 de doble propósito desde el punto de vista de la producción de carne y leche. El híbrido F1 resultante posee una inmunidad natural que le proporciona una resistencia a las garrapatas superior a la de Bos taurus, pero no tan eficiente como la de Bos indicus [4].
El mejoramiento de la resistencia adquirida de los bovinos contra la garrapata - logrado por cruzamiento genético- , tiene sus desventajas, debido a que la adquisición de resistencia no siempre está correlacionada con otras características deseables desde el punto de vista productivo. Además, la resistencia adquirida por esta vía usualmente es insuficiente como para controlar las infestaciones por el parásito, y, de esta forma, no logra evitar los grandes problemas que provocan en la ganadería [4, 5].
Naturaleza de la resistencia adquirida
Los estudios sobre el tema permiten presentar un mecanismo general para los eventos de adquisición y expresión de resistencia inmunológica contra las garrapatas, aunque obviamente éste puede variar de acuerdo con los diferentes hospederos y especies de garrapatas [6].
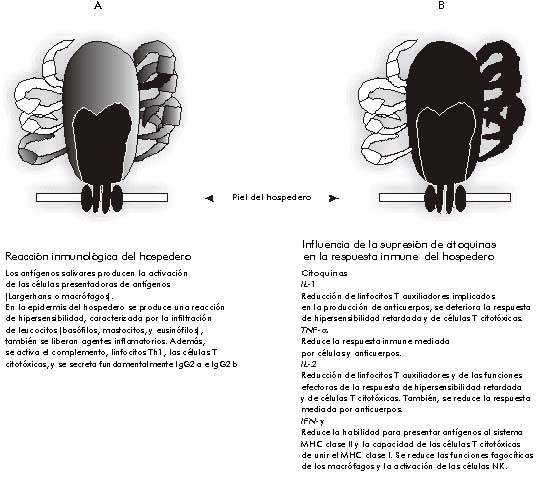
El comienzo de la alimentación de la garrapata sobre un animal que nunca antes estuvo expuesto a infestaciones por este parásito, se caracteriza por el reconocimiento de inmunógenos salivales por parte de las células de la epidermis y la dermis que se concentran en el sitio de la mordida. El tipo de inmunógeno introducido en el hospedero difiere durante el ciclo biológico del parásito. Proteínas y otras moléculas inmunogénicas presentes en la saliva de la garrapata, pueden ser procesadas por las células de Langerhans y por macrófagos en la epidermis y la dermis del animal. En los ganglios linfáticos, las moléculas salivales son procesadas por macrófagos o por células dendríticas, y, finalmente, son presentadas a linfocitos T específicos de antígenos en el contexto de los órganos linfoides secundarios. Así, los linfocitos T auxiliadores reconocen los inmunógenos de la garrapata en el contexto de las moléculas del complejo principal de histocompatibilidad (MHC, por sus siglas en inglés) clase II sobre la superficie de las células presentadoras de antígeno. Los linfocitos Th1 y Th2 son proveedores de señales inmunorreguladoras que permiten la generación de una respuesta mediada por células y anticuerpos (Figura 1) [7]. Además, los linfocitos Th1 son células que intervienen en las reacciones de hipersensibilidad tardía, incluida la respuesta de hipersensibilidad cutánea por infiltración de basófilos, que es generada por las garrapatas durante su alimentación (Figura 1) [8, 9]. Los inmunogénos, las células presentadoras de antígenos, los linfocitos T y las citoquinas contribuyen a la activación y diferenciación de los linfocitos B, los cuales producen anticuerpos circulantes homocitotrópicos y otros que reaccionan contra las garrapatas [10, 11]. Algunos estudios muestran que esta inmunidad disminuye en los períodos de preñez, lactancia y en condiciones de baja temperatura [12, 13]. La respuesta primaria de las garrapatas es inhibir las respuestas de hipersensibilidad y de anticuerpos generadas en el hospedero, pero se requiere información adicional para reconocer la naturaleza de los inmunogénos específicos que inducen, y subsecuentemente interactúan con, las defensas del hospedero contra las garrapatas.
La introducción de saliva de las garrapatas en la piel de un hospedero insensible, causa la desgranulación de mastocitos y basófilos, posiblemente mediante hidrólisis enzimática de la membrana plasmática producida por las enzimas salivales [14-16]. Por lo tanto, se liberan factores quimiotácticos y vasoactivos que pudieran contribuir a la formación de los modestos influjos de leucocitos observados en los sitios de unión de las garrapatas durante las primeras exposiciones del animal [17]. La generación de C5a por la activación de la vía alternativa del complemento, también pudiera contribuir a influjos celulares hacia el sitio donde pica la garrapata [18].
Las garrapatas pueden modular las respuestas innata y adquirida del hospedero, debido a que la saliva contiene inhibidores de la ruta alternativa de activación del complemento, de anafilotoxinas y de células asesinas naturales (NK, por sus siglas en inglés), los cuales son elementos de la defensa innata. Además, la saliva reduce la producción de citoquinas por los macrófagos, los cuales son muy importantes en los primeros pasos de desarrollo de una respuesta contra la garrapata. La alteración que ocurre se debe a que se afectan las señales requeridas para la estimulación de los linfocitos T y B. Por ejemplo, la capacidad de proliferación de los linfocitos T disminuye, al igual que la producción de IL-2 e IFN-g por los linfocitos Th1. Sin embargo, la producción de IL-4 por los linfocitos Th2 se mantiene normal (Figura 1) [19].
Figura 1. Caracterización de la inmunidad adquirida por los bovinos después de ser infestados con larvas de garrapata. A) Respuesta inicial del hospedero al enfrentarse con los antígenos salivales de las larvas. B) La respuesta inmunológica del hospedero es contrarrestada por sustancias secretadas por las larvas que se adhieren a la piel.
La especificidad de los anticuerpos inducidos por los inmunógenos salivales de la garrapata varía y aún está por definir su función. Además, una respuesta reducida de antígenos dependientes del timo pudiera ser el resultado de una función alterada de los macrófagos, los linfocitos T, o de otras interacciones no descritas aún.
Exposiciones repetidas o continuas provocan que las garrapatas que se alimentan del hospedero entren en contacto con los elementos de la repuesta inmune primaria. Por lo tanto, los inmunógenos salivales estimulan la respuesta de memoria de linfocitos T y B, los cuales garantizan una respuesta inmune potente después de la reinfestación con garrapatas. Es obvio definir que este tipo de defensa inmunológica está mediada por elementos característicos de la respuesta inmune adquirida. Además, es posible observar que durante el desarrollo de esta respuesta se presentan también elementos de la inmunidad innata, como las células de Langerhans, los macrófagos y las células dendríticas, que se mantienen como los elementos importantes en el procesamiento y presentación de antígenos, así como los mastocitos en la dermis y los basófilos cubiertos en su superficie por anticuerpos homocitotropos capaces de reaccionar con los inmunógenos introducidos en la saliva [20-22].
En animales resistentes, parece que los basófilos son atraídos por mediadores y linfocitos T al sitio de unión de la garrapata. Por ejemplo, la proteína C5a es un agente quimiotáctico generado por los anticuerpos circulantes que reaccionan con antígenos de la garrapata, lo que provoca la activación del complemento a través de la vía alternativa o la clásica. Además, los basófilos y los mastocitos se desgranulan cuando el complejo antígeno-anticuerpo homocitotropo se une a los receptores Fc de la superficie celular [23].
Los basófilos y la histamina liberada por los mastocitos, el leucotrieno B4 y los factores quimiotácticos de anafilaxis, atraen a los eosinófilos al sitio de la picada. Sin embargo, C5a es un factor quimiotáctico para los eosinófilos. Los neutrófilos son atraídos al sitio de unión de la garrapata por factores quimiotácticos de alto peso molecular derivados de basófilos y mastocitos. Los macrófagos que se han observado en las lesiones donde pican las garrapatas, probablemente están involucrados en la eliminación de antígenos, su procesamiento y la posterior presentación a los influjos de células inmunocompetentes.
Los mecanismos específicos que interrumpen la alimentación y reducen la ovoposición y la viabilidad de las garrapatas, aún se conocen poco en la actualidad. Las histaminas -proteínas básicas de los eosinófilos- , las prostaglandinas, las leucotrinas, las enzimas, y otras biomoléculas liberadas por los basófilos, los eosinófilos y los mastocitos, pueden, en conjunto, ser factores contribuyentes. Se conoce que las histaminas, los gránulos de los basófilos y los gránulos de los eosinófilos participan en la expresión de la resistencia adquirida [14]. Los anticuerpos, los linfocitos, el sistema del complemento y otros elementos de la respuesta inmune e inflamatoria participan en la resistencia adquirida, pero aún no se ha establecido su papel específico.
Concepto de antígenos ocultos
Los antígenos relacionados con la inmunidad adquirida para algunos parásitos, son un grupo de antígenos que están expuestos directamente al sistema inmune del hospedero. Estos antígenos normalmente coevolucionan hacia una adaptación durante la interacción hospedero-parásito, por lo que no debe ser sorprendente que la respuesta inmunológica del hospedero sea, cuando más, parcialmente protectora contra el parásito.
Durante los últimos años, se ha generalizado la idea de emplear nuevos antígenos para inducir la inmunidad del hospedero de forma artificial. Éstos son antígenos que no desempeñan ningún papel en la interacción hospedero-parásito. Por ejemplo, Schlein y Lewis [24], en 1976, vacunaron conejos con tejido muscular de la mosca Stomoxys calcitrans y observaron que éstas sufrían daño muscular después de alimentarse.
La primera sugerencia de utilización de los nuevos antígenos fue propuesta por Galun en 1975 [25], quien aportó la idea de que el animal puede ser inmunizado con las hormonas de desarrollo de la garrapata. Aún no hay reportes que avalen el éxito de la vacunación a partir de esta idea. Sin embargo, se ha observado un número de sistemas hospedero-garrapata en que se puede tener éxito a partir de la vacunación con extractos crudos de intestino de garrapatas o con el intestino íntegro [26]. Esta observación y la sugerencia anterior de Galun abren el camino de la investigación para obtener una vacuna contra B. microplus a través del empleo de los denominados antígenos ocultos, los cuales pueden ser clasificados como aquellas moléculas de notable importancia fisiológica para el parásito y que habitualmente no participan en la interacción hospedero-parásito.
Varios antígenos ocultos de B. microplus han sido aislados como candidatos vacunales, por ejemplo, Bm95 [27], Bm91 [28], QU13 [29, 30] y, el más conocido, Bm86 [31-35]. Otros antígenos ocultos son también la base del desarrollo de nuevas vacunas contra Lucilia cuprina, basadas en antígenos provenientes de la membrana peritrófica del insecto que, por su origen, fueron nombrados peritrofinas (PM95, PM48 y PM44) [36-38]. En la actualidad, hay varias vacunas en desarrollo contra otros ácaros basadas en antígenos ocultos [39], por ejemplo, contra Pediculus humanus [40], la mosca del gusano barrenador, Chysomya bezziana, y el piojo del salmón [40, 41].
Base inmunológica de la inmunidad inducida
Recientemente, se ha denominado inmunidad inducida a la inducción de una respuesta inmunológica protectora contra los parásitos basada en la inmunización del hospedero con antígenos provenientes de órganos y tejidos del parásito que normalmente no participan en las interacciones hospedero-parásito [42-45].
El efecto que tiene en las garrapatas la vacunación de bovinos con extractos antigénicos crudos, ha sido estudiado desde el punto de vista histológico. Se ha observado que las células digestivas se rompen y las células del hospedero penetran en la hemolinfa del parásito sin que se produzcan daños en los tejidos productores de saliva. Esta vacunación causó daños fundamentales en el estado adulto del parásito, pero que no fueron apreciables en el estado larval, donde ocurre sólo un retraimiento en su desarrollo a ninfa.
Un análisis de los componentes de la respuesta inmune de los bovinos vacunados, mostró que la IgG1, más que IgG2 e IgM, es responsable del daño provocado en las garrapatas. Aún no se tienen evidencias del papel de los linfocitos en la respuesta inmune. En un experimento in vitro, se observó que al introducir los linfocitos de animales vacunados en la sangre de bovinos controles (no vacunados), no se transmite ningún daño a las garrapatas [46]. Además, la adición de estos linfocitos a un animal inmune no aumenta apreciablemente el daño de las garrapatas [47].
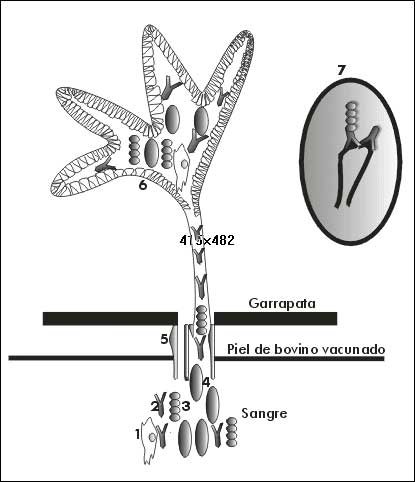
Actualmente, se conoce que el sistema del complemento desempeña un papel importante en la respuesta inmune que se genera contra el antígeno, y que es la causa de parte de los daños que se producen en el intestino de las garrapatas. Su papel fue determinado a partir de la observación del daño de las garrapatas que se alimentaban con sueros de animales vacunados, pero libres de toda actividad del complemento [48]. Las células del intestino que se ven afectadas esencialmente por la vacunación son las células digestivas, cuya función fundamental es endocitar y digerir intracelularmente la sangre como alimento fundamental del parásito (Figura 2) [49].
Figura 2. Mecanismo de acción de la inmunidad inducida por el antígeno oculto Bm86 en animales vacunados. Los elementos del sistema inmunológico presentes en la sangre, penetran a través del capítulo y la boca de la garrapata hasta llegar al intestino donde reaccionan con las moléculas blanco provocando la inhibición de la función biológica de las mismas. 1) Linfocitos; 2) anticuerpos anti-Bm86; 3) proteínas del complemento; 4) eritrocitos; 5) capítulo y boca; 6) intestino; 7) acción conjunta de anticuerpos anti-Bm86 y complemento sobre las células del intestino de la garrapata. (D) Moléculas de Bm86.
Los anticuerpos antiBm86 fueron capaces de reconocer sólo la membrana plasmática de las células digestivas de la garrapata (Figura 2), según se pudo observar en un experimento de inmunofluorescencia e inmunohistoquímica [50, 51]. En otros ensayos con sueros de animales vacunados con Bm86, se logró bloquear la endocitosis de albúmina conjugada con oro coloidal o fluoresceína in vitro [2] en ausencia de complemento [33]. Además, se determinó que la región Fab de los anticuerpos debe permanecer íntegra para la endocitosis [43, 50].
También se conoce que el ganado en estado de gestación es altamente susceptible a las infestaciones por garrapatas, debido fundamentalmente a su baja capacidad de respuesta inmunológica contra los antígenos presentados por la garrapata. Lo mismo ocurre en animales vacunados con el antígeno Bm86, debido a que el mayor por ciento de la inmunoglobulina de secreción IgG1 se encuentra en el calostro y la leche de estas vacas, lo que afecta sensiblemente la protección de los hatos vacunados [13].
Conclusiones
El desarrollo de una vacuna contra las garrapatas representa, en la actualidad, una de las opciones más prominentes para sustituir el control químico. Tiene la ventaja de ser específica de especie, inocua para el hombre y el medio ambiente, y fácil de administrar. El desarrollo notable de la inmunología molecular y la biotecnología, han permitido que se haga realidad el desarrollo de vacunas contra la garrapata. Sin embargo, el desarrollo de tales vacunas depende de la comprensión de la respuesta inmune adquirida del hospedero contra la garrapata.
Numerosos investigadores han utilizado homogenados totales de garrapatas y de glándulas salivales como inmunogénos vacunales, los cuales inducen niveles variables de protección. Estos investigadores seleccionaron inmunogénos lógicos basados en el conocimiento de la inmunidad del hospedero contra la garrapata; sin embargo, las vacunas más exitosas contienen antígenos definidos [44, 45]. Particular atención se ha prestado a los antígenos ocultos y a los inmunógenos del intestino [45, 52].
Estudios realizados con el antígeno oculto Bm86, resultaron en el desarrollo de una vacuna recombinante para limitar la infestación por la garrapata B. microplus [42, 52]. En resumen de todos los estudios realizados, se puede definir que el mecanismo de acción de esta vacuna contra B. microplus se basa en tres etapas que la hacen efectiva: 1) la unión de las inmunoglobulinas a las células digestivas o células blanco, 2) la acción del complemento por lo menos para amplificar el efecto de la unión de los anticuerpos, y 3) la inhibición de la endocitosis o función biológica de la molécula blanco como consecuencia de la unión de los anticuerpos.
Es necesario prestar una atención especial a la caracterización de las respuestas celular y humoral inducidas por la inmunización con estos antígenos, ya que las vacunas pudieran ser utilizadas en poblaciones de animales con una gran diversidad genética. En consecuencia, los inmunógenos seleccionados tienen que tener epítopos que sean procesados y presentados a las células inmunocompetentes en el contexto de la variedad de antígenos MHC. Se deberán utilizar sistemas de expresión que permitan obtener antígenos con características muy similares a las de la molécula natural. Finalmente debemos decir que durante la interacción hospedero-garrapata en un animal vacunado con antígenos ocultos, se desarrolla una inmunidad inducida que, junto a los elementos de la respuesta adquirida natural del hospedero, complementan una reacción de resistencia contra el parásito, el cual tendrá afectado de forma generalizada su ciclo biológico natural.
Agradecimientos
Agradezco al Dr. Eduardo Pentón su lectura crítica y las correcciones realizadas al documento.
References
Copyright Elfos Scientiae 2000
The following images related to this document are available:Photo images[ba00068b.jpg] [ba00068a.jpg] |
| |||||||||

{kind=link}
{kind=link}