 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnología Aplicada 2000;17:221-224 Obtaining Biologically Active IL-15 in Escherichia coli
@ Alicia Santos,1
Yanelys Morera,1 Manuel J Araña,1 Joel Ferrero,1
Alejandro Moro,1
1Centro de Ingeniería Genética y Biotecnología. Ave
31 e/ 158 y 190. AP 6162, CP 10600, Playa,
Received in December, 1999. Code Number: ba00069 Abstract Interleukin-15 (IL-15) is a recently discovered cytokine that shares with interleukin-2 (IL-2) the beta and gamma subunits of the receptor complex, therefore, both of them display similar biological effects. Moreover, IL-15 induces B and T cell proliferation, natural killer cell cytolytic activity and cytokine generation as well as protects T cell from apoptosis. The IL-15 synthesis and secretion are controlled both at transcriptional and post-transcriptional levels. This fine regulation poses a serious difficulty for its expression in eukaryotic systems. Here, the cDNA of human IL-15 was cloned into a prokaryotic vector under control of the tryptophan promoter for its expression in Escherichia coli. The recombinant IL-15 was obtained as an insoluble product, which represented 10% of the total protein. An extraction step in 8 M urea followed by a renaturalization process by diluting urea to 0.08 M was performed. The biological activity of this protein was 2.3 x 105 IU/mg in a CTLL-2 proliferation assay. The IL-15 proliferative activity on these cells was neutralized by a monoclonal antibody specific to IL-15, showing that the displayed activity was dependent on this cytokine. These simple and fast procedures allowed us to obtain a recombinant human IL-15 in E. coli, making this cytokine easily available for in vitro and therapeutic studies. Keywords: cytokine, eukaryotic system, expression, IL-15, recombinant E. coli Resumen Obtención de IL-15 biológicamente activa en Escherichia coli. La interleucina-15 (IL-15) es una citocina descrita recientemente que comparte con la IL-2 las subunidades beta y gamma del complejo receptor, y muestra muchos de los efectos biológicos descritos para la IL-2. La IL-15 induce la proliferación de células T, la maduración de células B, la citotoxicidad por células NK y la generación de citocinas, además de proteger a las células T de la apoptosis. La síntesis y secreción de la IL-15 es controlada a nivel transcripcional y post-transcripcional. Esta fuerte regulación representa una dificultad para su obtención biotecnológica en sistemas eucariontes. En este trabajo el ADNc de IL-15 humana fue clonado en un vector para su expresión en Escherichia coli bajo el control del promotor triptófano. La proteína fue expresada en forma insoluble y representó el 10% de la proteína total. Se realizó un proceso de extracción con 8 M de urea y de renaturalización por dilución hasta 0.08 M de urea. En el ensayo de proliferación con la línea celular CTLL-2 dependiente de citocinas, la proteína renaturalizada mostró una actividad específica de 2,3 x 105 UI/mg. La actividad proliferativa sobre estas células fue neutralizada mediante el uso de un anticuerpo monoclonal específico contra IL-15, muestra de que la actividad obtenida es dependiente de esta citocina. La obtención de la IL-15 humana recombinante en E. coli por un procedimiento sencillo permite disponer de esta citocina tanto para estudios in vitro como para su uso terapéutico. Palabras claves: citocina, E. coli recombinante, expresión, IL-15, sistema eucarionte Introduction Interleukin-15 (IL-15) is a member of the four a-helix bundle cytokine family. The effects of IL-15 are mediated by a trimeric membrane receptor comprising the b- and g-subunits of IL-2 receptor, and a different a-subunit [1]. Many biological effects of IL-2 and IL-15 are very similar. It has been shown that while IL-15 stimulates proliferation and cytolytic function of B, T, natural Killer (NK), and lymphokine-activated killer (LAK) cells, it is also a chemoattractant and protector from apoptosis of T cells [2–4]. Although several biological activities of IL-15 and IL-2 overlap, there are differences in the cellular sites of their synthesis and the molecular levels of regulation of their expression [5, 6] that might result in the preferential usage of one of these cytokines in a particular system. Although IL-15 mRNA is widely expressed in several normal tissues and tumors [7], its synthesis and secretion are controlled both at transcriptional and post-transcriptional levels [6]. Indeed, it has been shown that the 5’UTR of IL-15 messages are long, complex and contain several upstream AUGs, which drastically impair the efficiency of IL-15 translation. Moreover, IL-15 has an unusually long (48 aa) signal peptide, which could interfere with the efficient synthesis and secretion, as well as other elements present in the 3’ coding sequence that affect IL-15 production [8]. For these reasons, the expression of IL-15 without modification of its sequence is troublesome in eukaryotic systems. Here, the human IL-15 was expressed in the prokaryotic system Escherichia coli by cloning a cDNA that codes for a mature protein isolated from lipopolysaccharide (LPS)-stimulated peripheral blood mononuclear cells (PBMC), and after an extraction and a renaturalization process, a biological active protein able to stimulate proliferation of the cytokine-dependent murine T-cell line CTLL-2, was obtained with more than 90% purity. A neutralizing monoclonal antibody (MAb) to human IL-15 completely inhibited the proliferative activity induced by this bacterially expressed IL-15. Materials and Methods Monocyte preparation PBMCs from healthy volunteers were obtained by Ficoll-Hypaque (Pharmacia, UK) gradient centrifugation after informed consent was received. Monocytes were purified by Percoll gradient. Cells were cultured in RPMI 1640 (Gibco-BRL, USA) containing 2 mM L-glutamine and 10% heat inactivated fetal bovine serum, in presence of 5 mg/mL of LPS, for 6 h at 37 ºC and 5% CO2. Reverse transcription-polymerase chain reaction (RT-PCR) Total RNA was purified with TriReagent (Sigma, USA) as specified by the manufacturer’s instructions. cDNA synthesis was performed with 1 mg of total RNA from activated monocyte in a 20-mL volume reaction, containing 50 mmol/L Tris-HCl (pH 8.3), 75 mmol/L KCl, 3 mmol/L MgCl2, 40 U of RNase inhibitor, 0.1 mg oligo (dT), and 20 U AMV reverse transcriptase (Promega, USA). The reaction mixture was incubated at 42 ºC for 1 h and stopped at 90 ºC for 5 min. A 3-mL aliquot of the obtained cDNA was amplified by PCR in a 50-mL reaction: 500 mmol/L KCl, 100 mmol/L Tris-HCl buffer (pH 8.8), 25 mmol/L MgCl2, 2 mmol/L of each dNTP, 25 pmol of each primer, and 5 U Taq DNA polymerase (Enzibiot, Cuba). The following oligonucleotides were used in the PCRs: IL-15 sense 5’CATGCCATGGCAAACT GGGTGAATGTAATAAGTGAT3’, and IL-15 antisense, 5’CGGGATCCCGTTAAGAAGTGTT GATGAACAT3’. These oligonucleotides contained NcoI-BamHI restriction sites for easier cloning. Cycling conditions: 1 min at 94 ºC, 1 min at 58 ºC and 1 min at 72 ºC, for 30 cycles. PCR was performed on a DNA Thermal Cycler (MJ Research, USA) and the reaction was subjected to electrophoresis on type I-A: low EEO agarose (Sigma, USA). DNA was extracted according to standard phenol-chloroform protocol [9]. Bacterial strains
W3110: F- thyA36 deoc2 IN(rrnD-rrnE)1 rph pyrE lambda- TG-1: F’ traD36 proAB lac§q ZDM15 thi D(lac-proAB) supE hsdS M-15: F- NaIs Strs rifs lac- ara- gal- mtl- recA+ uvr+ MC1061: F- araD139 D(ara-leu)7696 galU galK D(lac)X74 rspL (Strr) thi
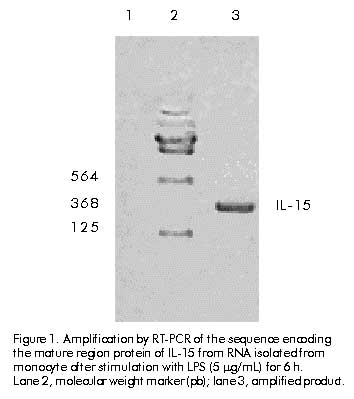
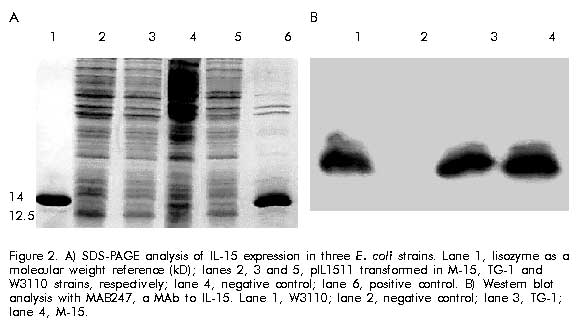
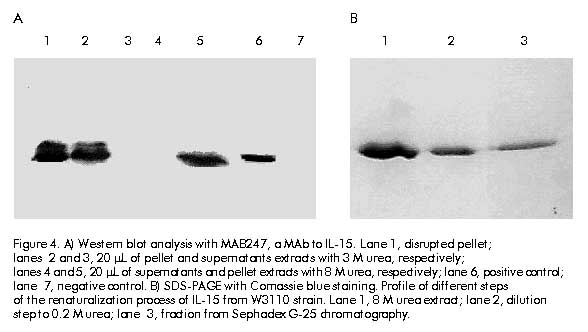
Cloning and expression The amplified gene was ligated into the vector pIL-2 [10] digested with NcoI-BamHI restriction enzyme (Promega, USA) replacing IL-2 gene, under the control of the tryptophan promoter, using ampicillin resistance gene. Ligations were transformed into E. coli strain MC1061 [11]. The recombinant clones were selected by restriction analysis and DNA sequencing [12]. The clone selected was named pIL1511. E. coli strains W3110, TG-1 and M-15 containing the pIL1511 vector were grown overnight at 37 ºC in 5 mL Luria broth (LB) medium (tryptone 10 g/L, yeast extract 5 g/L, NaCl 10 g/L). Later, 500 mL were taken to inoculate 50 mL of enriched M9 medium (NaHPO4 6 g/L, KH2PO4 3 g/L, NaCl 0.5 g/L, NH4Cl 1 g/L, MgSO4 1 mM, CaCl2 10 mM, glucose 0.2%, casein hydrolysate 2%) and further incubated for 2 h at 37 ºC. Then, the induction was performed during 12 h at 37 ºC in presence of 3b-indolacrylic acid (40 mg/mL) for 12 h. Antibodies and cytokine Human recombinant IL-15 (WHO Standard Reference Number 95/554, NIBSC, UK) was used as international standard (10 000 IU/mL), and the MAb to IL-15 (MAB247, IgG1, 0.1 mg/mL; R&D, USA), as a neutralizing antibody in the proliferative assay. Human recombinant IL-15 (20 ng/mL; R&D, USA) was used for Western blot analysis. Western blot analysis For Western blot analysis a 15% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was resolved with 20 mg of each sample per well, and transferred into Hybond-C membrane in a semi-dry transfer system (Bio-Rad, USA) [13]. The membrane with the transferred proteins was incubated overnight at 4 ºC with phosphate-buffered saline (PBS, 0.1 M NaCl, 2 mM KCl, 10 mM Na2HPO4, 1 mM KH2PO4, pH 7.2) containing 5% skim milk. The membrane was incubated using anti-IL-15 MAb (MAB247) (1 mg/mL) as the primary antibody for 1 h at 37 ºC, then washed several times (5 min each) with PBS. Specific antibodies bound to protein bands were detected with an anti-mouse peroxidase conjugate diluted 1:2000 by enhanced chemoluminescence (ECL, Amersham Pharmacia Biotech, UK) according to manufacturer’s instructions. Twenty micrograms of protein from E. coli W3110 strain, transformed with pIL-2 induced in the same conditions as pIL1511 were used as negative control, and recombinant human IL-15 from R&D was used as positive control. Protein purification Cells were collected after 12 h of induction, and disrupted using an ultrasonic disrupter (Braun, Germany). Taking advantage of the insolubility of the IL-15 recombinant protein, it was purified by washing the cellular pellet with TE buffer (Tris 0.1 M, EDTA 0.01 M, pH 8) and 3 M urea as a chaotropic agent [14]. The partially purified protein was further solubilized using 8 M urea in TE buffer. For the renaturalization process the chaotropic agent was removed by quickly diluting the protein solution to 0.08 M urea and passing it through Sephadex G-25 (Pharmacia, Sweden) chromatography in Tris buffer (100 mM, NaCl 10 mM, pH 7.6). Total protein concentration in the samples was determined by the method of Bradford [15] using bovine serum albumin as standard. The expression level and purity were determined by densitometry using the Molecular Analyst Software (Bio-Rad, USA). Proliferation assay CTLL-2 is a cytokine-dependent murine T-cell line which proliferates in response to human IL-15 [6]. The bioassay was performed in a 96-well cell culture cluster dish (Costar, USA) at 5 x 103 CTLL-2 cells per well, previously washed three times with medium without serum, and incubated in presence of twofold serial dilutions (starting dilution 1:1000) of recombinant human IL-15 international standard and the IL-15 obtained from the renaturalization process in 100 mL total volume at 37 ºC and 5% CO2. After 48 h 1-[4,5-dimethylthiasol-2-g1]-3,5 diphenylformazan (MTT) [16] was added and the plates were further incubated for 4 h. Finally, 50 mL of a solution containing 10% SDS, 0.1N HCL and 50% isopropylalcohol were added per well. Plates were read at 570 nm on a Multiscan (Sensident Scan, Merck, Germany). Regression, parallelism and lineality of the results were analyzed by the exact Fisher’s test. The specific activity was calculated by the ratio of activity determined by CTLL-2 assay and the protein concentration of recombinant product of Sephadex G-25 chromatography. Results and Discussion To clone the mature region of human IL-15, cDNA from LPS-induced PBMC was amplified by PCR with specific primers containing the end restriction site, the methionine codon as initiation of translation site and the stop codon. The amplified product was purified from gel and cloned into the pIL-2 vector, under control of the tryptophan promoter (Figure 1). Figure 1. Amplification by RT-PCR of the sequence encoding the mature region protein of IL-15 from RNA isolated from monocyte after stimulation with LPS (5 mg/mL) for 6 h. Lane 2, molecular weight marker (pb); lane 3, amplified product. To compare the expression levels, the IL-15 gene was expressed in three different E. coli strains (W3110, TG-1 and M-15). Cells induced during 12 h were collected and analyzed by SDS-PAGE. Similar expression levels (about 10% of total cellular protein) were observed in all strains (Figure 2A). The expression of the IL-15 heterologous product was confirmed by immunodetection using a specific MAb (Figure 2B). Figure 2. A) SDS-PAGE analysis of IL-15 expression in three E. coli strains. Lane 1, lisozyme as a molecular weight reference (kD); lanes 2, 3 and 5, pIL1511 transformed in M-15, TG-1 and W3110 strains, respectively; lane 4, negative control; lane 6, positive control. B) Western blot analysis with MAB247, a MAb to IL-15. Lane 1, W3110; lane 2, negative control; lane 3, TG-1; lane 4, M-15. Cells were disrupted and IL-15 protein was found in the insoluble fraction (Figure 3). The cellular pellet was consecutively washed with TE and 3 M urea, and the protein was completely extracted in 8 M urea. Protein yield was not affected after washing (Figure 4A). After a renaturalization process, a single band at 12.5 kD using SDS-PAGE followed by Coomassie blue staining and densitometry in gel electrophoresis, confirmed that IL-15 was the dominant protein in the preparation with 90% purity (Figure 4B). Figure 3. Western blot analysis of cellular localization. Lanes 1 and 2, negative controls; lane 3, total protein; lane 4, disrupted pellet; lane 5, disrupted supernatant. Figure 4. A) Western blot analysis with MAB247, a MAb to IL-15. Lane 1, disrupted pellet; lanes 2 and 3, 20 mL of pellet and supernatants extracts with 3 M urea, respectively; lanes 4 and 5, 20 mL of supernatants and pellet extracts with 8 M urea, respectively; lane 6, positive control; lane 7, negative control. B) SDS-PAGE with Comassie blue staining. Profile of different steps of the renaturalization process of IL-15 from W3110 strain. Lane 1, 8 M urea extract; lane 2, dilution step to 0.2 M urea; lane 3, fraction from Sephadex G-25 chromatography. Using as a reference an international standard of IL-15, this fraction was tested on CTLL-2 cell line in order to determine the proliferative activity. As shown in Figure 5, the IL-15 obtained using this procedure induces the proliferation of CTLL-2 cell line in a dose-dependent manner with an specific activity of 2.3 x 105 IU/mg. Fisher’s exact statistical test showed lineality and parallelism (P > 0.05), and the regression was highly significant (P < 0.01). Therefore, during the renaturalization process IL-15 is obtained in a correct functional conformation. Figure 5. Induction of CTLL-2 proliferation by IL-15 preparation. CTLL-2 cells were incubated with serial dilutions of an international standard of IL-15 and IL-15 obtained from E. coli initially diluted to 103 and in presence of the neutralizing monoclonal antibody to human IL-15, during 48 h at 37 ºC, and 5% CO2. Proliferation was measured by MTT colorimetric assay. Fisher’s exact test showed significant levels of 0.0001, 0.3150, and 0.2953 for regression, parallelism and lineality, respectively. To demonstrate the specificity of the biological effect in the extract, a neutralizing MAb to human IL-15 at 0.1 mg/mL was used, and it was observed that the proliferative response was totally inhibited in presence of this antibody. Thus, it was demonstrated the feasibility of the E. coli bacterial system to express a complex eukaryotic protein, such as IL-15, in its biologically active form. No toxic effects were observed in bacteria, at least 12 h after induction. Finally, the addition of an N-terminal methionine and the lack of glycosylation, two of the principal disadvantages of this system [17], did not affect the in vitro biological activity of the IL-15 expressed in E. coli. References
Copyright Elfos Scientiae 2000
The following images related to this document are available:Photo images[ba00069c.jpg] [ba00069f.jpg] [ba00069d.jpg] [ba00069a.jpg] [ba00069b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}