 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnología Aplicada 2000;17:241-246
Estudio de la variabilidad inducida en células y plántulas de cebolla (Allium cepa, L.) cv. Caribe-71 regeneradas in vitro @ Amelia Capote Rodríguez, Zoila Fundora Mayor, Odalys Pérez Díaz Instituto de Investigaciones Fundamentales en Agricultura Tropical
"Alejandro de Humboldt" (INIFAT).
Recibido en marzo del 2000. Code Number: ba00073 RESUMEN Se estudió el efecto de diferentes reguladores del crecimiento sobre la inducción y crecimiento de callos a partir de segmentos de hojas inmaduras, y la regeneración de plantas a partir de éstos. También se evaluaron, mediante estudios citogenéticos y bioquímicos (isoenzimas peroxidasas), las posibles variaciones que se pudieran presentar durante el período de cultivo in vitro. Los resultados obtenidos muestran que, en las células presentes en los callos, se observan diferentes niveles de ploidía en dependencia del tipo y concentración de los reguladores adicionados. Las plantas regeneradas a partir de callos muestran diferencias en algunos caracteres morfológicos relacionados con los rendimientos, pero mantienen su condición diploide, lo que evidencia la existencia de un proceso de selección en la diferenciación celular. Los patrones isoenzimáticos muestran variaciones en los callos y las plántulas regeneradas en relación con el material original. Se sugiere utilizar estos resultados en investigaciones encaminadas a la aplicación de las técnicas biotecnológicas a la selección de una nueva variabilidad genética, y profundizar en estos estudios mediante el empleo de otras técnicas moleculares que permitan hacer un análisis más detallado del genoma de estas plantas.
Palabras claves: Allium cepa, cebolla, inestabilidad genética, variación somaclonal
ABSTRACT Study of the Variability Induced in In Vitro Cells and Regenerated Plants of Onion (Allium cepa, L.) cv. Caribe-71. The effect of different growth regulators on callus induction and growth from segments of young leaves, was studied. Plant regeneration from these calli was also studied. Variations that might be present during the in vitro culture period were evaluated by means of cytogenetic and biochemical (peroxidase patterns) studies. The results showed that callus cells have different ploidy levels depending on the type and concentration of the regulators added. Plants regenerated from calli showed differences in their morphological traits but kept their diploid condition, which evidenced the existence of a selection process during cell differentiation. Peroxidase patterns showed variations in the calli and the regenerated plants in relation to the original material. We suggest the use of these results in research works concerning the application of biotechnological techniques to obtain a new genetic variability, as well as the continuation of these studies using other molecular techniques that enable us to gain insights into the genome of these plants. Keywords: Allium cepa, genetic instability, onion, somaclonal variation Introducción La cebolla (Allium cepa, L.) es una especie hortícola de gran importancia económica para Cuba. Sin embargo, debido a las dificultades que existen para su cultivo, se realizan numerosos esfuerzos con el objetivo de buscar nuevas variedades que se adapten mejor a las condiciones climáticas de ese país caribeño, y que garanticen rendimientos más elevados. El mejoramiento genético de este cultivo está limitado cuando se usan las técnicas tradicionales, debido a que no todos los cultivares producen semillas y a la existencia de barreras de incompatibilidad con las variedades más rústicas, portadoras de los genes de resistencia a enfermedades y de adaptación ambiental [1]. Resulta de gran interés para el mejoramiento genético, el descubrimiento de la alta frecuencia de células cultivadas in vitro que muestran mutaciones que se pueden expresar en las plantas regeneradas de forma adventicia [2], como es el caso de la cebolla [3]. Por esta razón, el cultivo de tejidos y células in vitro constituye una alternativa en los programas de mejoramiento genético, puesto que durante este período se generan modificaciones genéticas heredables denominadas variaciones somaclonales [4]. Estas variaciones posibilitan la obtención de una nueva variabilidad genética [5]. Se ha planteado que estas variaciones dependen de condiciones preexistentes en el explante inicial y/o de factores externos como la composición de los medios de cultivo, sobre todo el tipo y la concentración de los reguladores del crecimiento adicionados [6]. Se ha observado la variación en el número de cromosomas en los cultivos in vitro [7]. No obstante, se ha estudiado poco el cariotipo de las células en el momento de su cultivo, su cariotipo posterior como células libres o callos, y las plántulas producidas por éstos [8]. El objetivo de este trabajo fue estudiar el efecto de los reguladores del crecimiento en la inducción de callos y la regeneración de plantas, así como estudiar las posibles variaciones que se pueden encontrar en las células y las plántulas de cebolla cultivadas in vitro. Materiales y métodos Material vegetal Para desarrollar este trabajo, se seleccionó el cv. Caribe-71 por ser la variedad comercial de mayor distribución en el país, debido a su adaptación a las condiciones climáticas y a sus rendimientos aceptables. Obtención de callos Se utilizaron como explantes segmentos de hojas inmaduras, los cuales fueron lavados con agua corriente y detergente comercial. Posteriormente y en condiciones asépticas, fueron sumergidos en etanol 70% durante 10 min y colocados en una solución de hipoclorito de sodio 5% durante 20 min. Una vez concluida la desinfección, se lavaron tres veces en agua destilada estéril y se cultivaron en medio MS [9] suplido con sacarosa 30 g/L y diferentes combinaciones de auxinas y citoquininas (Tabla 1). En todos los medios de cultivo utilizados, se ajustó el pH a 5,8 en un pH metro digital modelo MV 780 (PRÄCITRONIC, Alemania), antes de añadir el agar (7 g/L). Los medios de cultivo se esterilizaron en un autoclave a 121 ºC y 1,2 atm de presión, durante 20 min. Cada tratamiento tuvo 24 réplicas. Los cultivos fueron mantenidos en la oscuridad por un período de 90 días a una temperatura de 25 ± 2 ºC. Durante ese tiempo, se realizaron subcultivos en el mismo medio de cultivo cada 30 días, con el propósito de lograr su establecimiento óptimo. El crecimiento relativo de los callos se determinó después de los 90 días de cultivo mediante la evaluación del incremento de peso en relación con el peso inicial de los explantes (aproximadamente 0,05 g), y se expresó en por ciento según la siguiente escala de valores: 0, no crecimiento (0%); 1, aumento del tamaño del explante (1%-25%); 2, poco crecimiento (26%-50%); 3, crecimiento medio (51%-75%); y 4, crecimiento abundante (76% o más). Regeneración de plantas Los callos obtenidos en el medio de mayor crecimiento (MS + picloram 0,1 mg/L y 6g, g-dimetilalilaminopurina [2iP] 1 mg/L) fueron trasplantados al mismo medio mineral suplido con diferentes reguladores del crecimiento (Tabla 1), con el objetivo de inducir la regeneración de brotes. Tabla 1. Composición de los medios de cultivo empleados para la obtención de callos y para la regeneración de plantas de cebolla a partir de éstos.
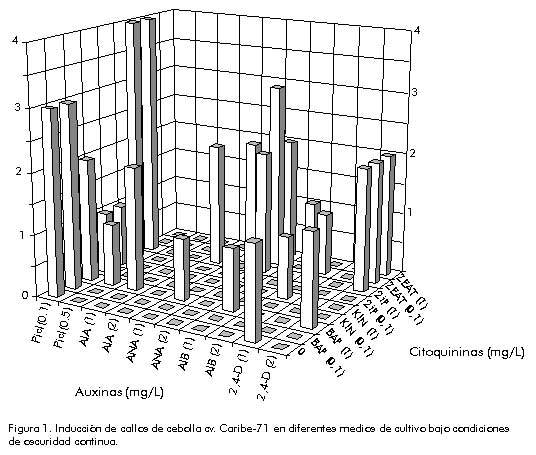
2,4-D, ácido 2,4 diclorofenoxiacético; AIB, ácido indolbutírico; ANA, ácido naftalenacético; KIN, 6-furfurilaminopurina; BAP, 6-bencilaminopurina; 2iP, 6-g , g -dimetilalilaminopurina; picloram, ácido 4-amino 3,5,6-tripicolínico. Los cultivos fueron mantenidos en un fotoperíodo de 14 h luz a una intensidad luminosa de 2 000 lux y a una temperatura de 25 ± 2 ºC, por un período de 90 días, tiempo después del cual se evaluó la aparición de brotes, zonas nodulares verdes y raíces en un total de 15 réplicas. Estudio de las variaciones inducidas en células y plantas regeneradas in vitro Estudios citogenéticos. Se realizaron diferentes estudios con los callos y las plantas regeneradas in vitro, con el propósito de determinar los cambios en los niveles de ploidía. Los callos de 90 días fueron tratados previamente con una solución de 8-hidroxiquinolina 0,002 M durante 4 h a temperatura ambiente, fijados en etanol-ácido acético (3:1) e hidrolizados con HCl 1 N. El aplastado se realizó en orceína acética y se contó el número de células en metafase y el número de cromosomas por célula. Se evaluó un total de 10 preparaciones por variante. Se estudió, además, el número de cromosomas de las células de los ápices radiculares de las plantas regeneradas in vitro a partir de callos, con el propósito de determinar el nivel de ploidía de las plantas obtenidas. Se evaluó un total de 50 plántulas seleccionadas al azar. Estudio de los caracteres agromorfológicos. El estudio de los caracteres agromorfológicos se realizó con el objetivo de detectar las posibles desviaciones fenotípicas de las plantas regeneradas. Para ello, se estudiaron diferentes caracteres en dos grupos de plantas: plantas regeneradas in vitro y plantas obtenidas a partir de semillas del cultivar original. Ambos grupos de plantas fueron sembrados en iguales condiciones y con las mismas atenciones culturales. Se evaluaron los siguientes caracteres: sabor, color externo e interno del bulbo, peso, diámetro y altura del bulbo, número de yemas y número de catafilos. Se analizó un total de 35 plantas por variante y los resultados individuales se evaluaron estadísticamente mediante un análisis factorial discriminante, según el programa estadístico STAT-ITCF con el empleo de la distancia generalizada de Mahalanobis. Estudio de los patrones isoenzimáticos. Para los análisis electroforéticos de las isoperoxidasas, se utilizaron las hojas de las plántulas germinadas in vitro a partir de semillas (plantas controles), los callos crecidos en el medio de mayor crecimiento (MS + 0,1 mg/L picloram y 1 mg/L 2iP) y las hojas de plántulas regeneradas in vitro (MS + 2,4 D 0,01 mg/L y KIN 5 mg/L). En todos los casos se utilizó material vegetal de cuatro semanas de edad. La extracción se realizó macerando el material vegetal (2 g) en tampón fosfato 0,1 M con sacarosa 10%, pH 7,0, en una relación de 1 mL de tampón por cada gramo de tejido para las hojas (1:1 v/p) y 2 mL de tampón por cada gramo para los callos (1:2 v/p). Las corridas electroforéticas se realizaron en geles de poliacrilamida 5%. Se aplicaron 10 mL de muestra por pocillo. Se empleó un sistema electroforético horizontal que permite la separación simultánea de las bandas aniónicas y catiónicas. El tiempo de corrida fue aproximadamente de 5 h con una intensidad de corriente de 1 mA por pocillo. Tanto la extracción como la electroforesis se llevaron a cabo a una temperatura de 4 ºC. La tinción se realizó en una solución de bencidina 25% y peróxido de hidrógeno 0,01%, y los zimogramas se confeccionaron a partir de la medición de la posición de la bandas con respecto al punto de aplicación (movilidad), y de su intensidad. Resultados y Discusión Obtención de callos y regeneración de las plantas Cuando se analizó la respuesta en el crecimiento según la composición de los diferentes reguladores en los medios de cultivo (Figura 1), se observó que la adición de 2,4-D 2 mg/L estimuló la formación de callos, siempre que estuvo en presencia de 2iP o zeatina. Sin embargo, en general, el crecimiento de los callos fue deficiente en presencia de esta auxina. Figura 1. Inducción de callos de cebolla cv. Caribe-71 en diferentes medios de cultivo bajo condiciones de oscuridad continua. Con la adición de ácido naftalenacético (ANA), se obtuvo una respuesta similar a la del 2,4-D, pero se indujo la regeneración de raíces en los callos. Esto confirma que esta auxina tiene una excelente capacidad de inducir la rizogénesis in vitro [10], aspecto que no es deseado cuando se quiere lograr un crecimiento óptimo de los callos. Las mejores respuestas se obtuvieron con la adición de picloram a baja concentración (0,1 mg/L). Esta auxina resultó ser la más efectiva para inducir un crecimiento óptimo de los callos, el cual se estimula con la adición de 2iP al medio. Se obtuvieron, además, callos altamente friables y de coloración cremosa, características que están asociadas a una mayor capacidad de regeneración de plantas. Se plantea que la eficiencia del picloram en la inducción de callos es comparable a la del 2,4-D en cuanto a la inducción del crecimiento, y superior a éste en cuanto a lograr la friabilidad de los callos y la subsiguiente regeneración de plantas de Allium cepa [11] y Allium trifoliatum subsp. hirsutum [12]. El picloram es una auxina sintética muy eficiente para el cultivo de tejidos vegetales, puesto que ofrece ciertas ventajas en relación con el 2,4-D por ser soluble en agua, menos tóxica por su efectividad a concentraciones más bajas y, en último lugar, por ofrecer un mayor potencial para la regeneración de plantas a partir de callos [13]. En cuanto a las citoquininas empleadas, el 2iP y el BAP estimulan, en mayor medida, el crecimiento de los callos cuando están en presencia de auxinas en el medio de cultivo. En la Tabla 2 se muestra la aparición de brotes, zonas nodulares verdes y raíces según los diferentes medios de cultivo utilizados. La respuesta obtenida está asociada al tipo de regulador del crecimiento adicionado. Tabla 2. Aparición de brotes, zonas nodulares verdes y raíces para los diferentes medios de organogénesis utilizados.
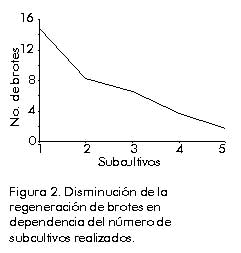
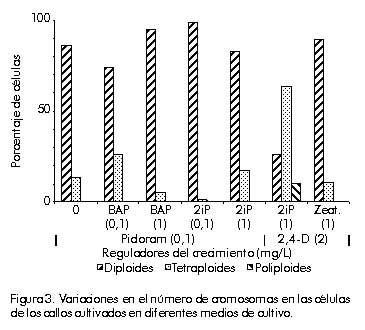
*(1-5 ); **(5-10); ***(más de 10) La diferenciación de brotes ocurrió en los medios suplidos con KIN y BAP como citoquininas, aunque con menor frecuencia. Por otra parte, la adición de 2iP no fue efectiva debido a que no estimuló la formación de brotes ni de zonas nodulares verdes. La formación de estas zonas fue estimulada por la presencia de ANA; sin embargo, no fue posible su desarrollo posterior a brotes. Según los resultados obtenidos, la adición de KIN a concentraciones altas (2 y 5 mg/L), en combinación con 2,4-D a una concentración muy baja (0,01 mg/L), genera el medio más eficiente para la regeneración de plantas. Sin embargo, aún en los medios de mejor comportamiento, el número de brotes regenerados es bajo (10 a 15 brotes/callo). Se detectó, además, que la capacidad de regeneración disminuye a medida que aumenta el número de subcultivos (Figura 2), lo que pudiera deberse a que, después de cierto período de cultivo, el potencial morfogenético del callo disminuye. Este hecho está asociado a cambios genéticos que se producen en las células en cultivo, puesto que esta variación es influida, entre otros factores, por la duración del período de cultivo [14]. Figura 2. Disminución de la regeneración de brotes en dependencia del número de subcultivos realizados. Eady et al. [15] demostraron que se pueden obtener callos de cebolla altamente embriogénicos con la adición de 2,4-D y picloram al medio de cultivo. Sin embargo, una característica de los protocolos publicados hasta el momento para esta especie, es el número relativamente bajo de brotes formados independientemente del explante utilizado, lo cual ha motivado, entre otras cosas, la ausencia de un protocolo de transformación genética estable para este cultivo [16]. La regeneración eficiente de callos de monocotiledóneas ha mostrado ser dependiente de varios factores, como el tipo de explante, el genotipo, el medio de cultivo y el tipo de callo obtenido [17]. En el caso específico de la cebolla, se plantea que la regeneración de brotes depende de numerosos factores, de los cuales el más importante es el nivel de reguladores del crecimiento presentes en el medio de cultivo. Para este proceso se necesitan concentraciones más altas de citoquininas que de auxinas [18]. Estudios de la variación inducida en los callos y las plantas regeneradas Estudios de los caracteres citogenéticos. Las células presentes en los callos provenientes de medios de inducción, muestran una alteración en los niveles de ploidía en dependencia del tipo de reguladores adicionados (Figura 3). Figura 3. Variaciones en el número de cromosomas en las células de los callos cultivados en diferentes medios de cultivo. En los callos obtenidos en medios de cultivo con 2,4-D como auxina, se obtuvo un gran número de células tetraploides (2n = 2x = 32 cromosomas), poliploides (más de 60 cromosomas) y células con hasta 12 nucléolos, lo que indica que, como mínimo, triplicaron sus cromosomas. La presencia de estas células aumentó cuando se combinó con el 2iP. Entre los reguladores del crecimiento, el 2,4-D . auxina sintética utilizada comúnmente en los medios de cultivo. ha demostrado tener un gran efecto mutagénico en las células meristemáticas, las cuales sufren cambios sustanciales en la expresión génica [19]. Se ha planteado que el 2,4-D estimula generalmente la síntesis de ADN, lo que puede conducir a un proceso de endorreplicación y, por otra parte, reduce el índice mitótico y aumenta el sobrecruzamiento mitótico. Los dos últimos fenómenos incrementan la poliploidía en las células cultivadas [20]. Por otra parte, las hormonas involucradas en la senescencia celular, entre ellas las citoquininas, desempeñan un papel importante en la variación inducida, debido a que el proceso de senescencia está asociado con la estabilidad del genoma [7]. Ésta pudiera ser la causa de la pérdida del potencial organogenético después de un período prolongado de cultivo, puesto que esta pérdida está asociada a una acumulación de cambios genéticos que ocurren en las células en cultivo [21]. Sin embargo, en los callos crecidos en medios suplidos con picloram, se encontró que la mayor cantidad de células mantienen su condición diploide (2n = 2x = 16 cromosomas). La adición de BAP 0,1 mg/L fue lo que estimuló la aparición de células tetraploides en un mayor por ciento (25,93%). No hay duda de que las aberraciones cromosómicas, especialmente los cambios en los niveles de ploidía, son comunes en las células y los tejidos cultivados en condiciones asépticas [22]. Aún no se conoce bien el modo de acción de los reguladores del crecimiento sobre los cambios en los niveles de ploidía. Según Bayliss [23], no hay un efecto directo de los componentes del medio que lleve a una inestabilidad cromosómica, sino que ésta resulta del crecimiento desorganizado de los callos, que es controlado por los reguladores del crecimiento. El balance hormonal es crítico para el crecimiento celular y la estabilidad del genoma. Se ha demostrado que el proceso de desdiferenciación celular está acompañado de un cambio en los niveles de metilación [24], así como de una variación en la talla del genoma por amplificación o disminución de las secuencias repetitivas [25]. También se plantea que la aparición y el grado de ploidía obtenido in vitro, puede ser el resultado de un equilibrio entre la constitución genética de la especie y la composición del medio de cultivo. Estos resultados indican que la presión de selección del medio de cultivo puede llevar a la manifestación del control genético en el comportamiento de los cromosomas, de una manera diferencial según la especie [26]. Se ha observado que el cariotipo varía en las células cultivadas, las plantas regeneradas y su progenie en numerosas especies, incluidas variaciones en el reordenamiento de los cromosomas como los cambios en los niveles de euploidía y aneuploidía. Esta última afecta la capacidad de las células para regenerar plantas, particularmente en las especies diploides [7]. A nivel molecular, estas mutaciones están relacionadas frecuentemente con los procesos de endorreduplicación o amplificación del ADN. Ambos procesos traen como consecuencia el aumento del contenido de ADN que se puede producir durante los procesos morfogenéticos, o bien en condiciones de estrés [27]. En los últimos años, ha surgido una teoría que considera al genoma vegetal en un estado de flujo constante, que se puede producir entre los ciclos mitótico o meiótico. Esta teoría postula que los mecanismos por medio de los cuales se producen estas variaciones, incrementan mil veces su frecuencia de aparición en condiciones de cultivo de tejidos. Esta fluidez permitiría la modulación rápida del genoma frente a situaciones de estrés y sería el mecanismo por medio del cual los vegetales se adaptan a diferentes condiciones. Por otra parte, la variación somaclonal aparece como consecuencia de este fenómeno [22]. Estudio de los caracteres morfológicos. Al analizar los resultados obtenidos cuando se evaluaron los caracteres morfológicos de las plantas regeneradas y del cultivar original, se observó que la función discriminante entre ambos grupos indica que las vitroplantas difieren significativamente de las plantas del cultivar original (Tabla 3). Tabla 3. Resultados del análisis discriminante realizado
para los caracteres morfológicos
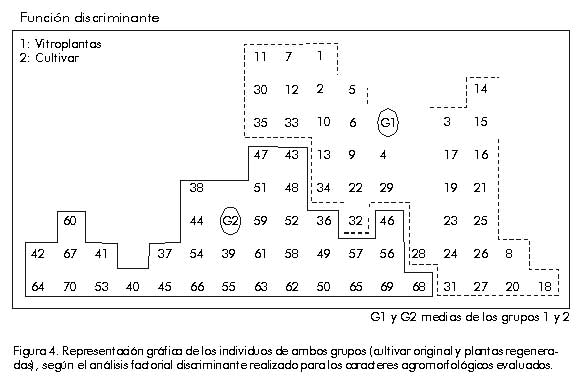
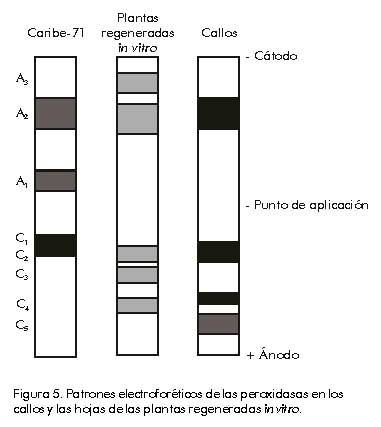
DE, desviación estándar. Los valores subrayados corresponden a las variables que más contribuyen a la diferencia entre grupos. El por ciento de individuos bien calificados fue 74,6% y las mayores diferencias entre ambos grupos de plantas se detectaron fundamentalmente en el peso y diámetro de los frutos, parámetros que están estrechamente vinculados entre sí y determinan el rendimiento en este cultivo [28]. Si se analiza el gráfico correspondiente a la distribución de los individuos de acuerdo con los valores de sus funciones discriminantes (Figura 4), se puede apreciar que existe una dispersión muy grande del valor de las funciones dentro de cada grupo con respecto a los valores promedios de cada uno (G1 y G2), lo que indica una variación en los caracteres estudiados. Figura 4. Representación gráfica de los individuos de ambos grupos (cultivar original y plantas regeneradas), según el análisis factorial discriminante realizado para los caracteres agromorfológicos evaluados. Por otra parte, los caracteres cualitativos evaluados, como color externo e interno del bulbo y su sabor, se mantuvieron estables para los dos grupos de plantas analizadas. Sin embargo, cuando se realizó el análisis de los cromosomas en los ápices radiculares de las plántulas regeneradas a partir de callos, se observó que éstas mantuvieron su condición diploide. Estos resultados coinciden con los obtenidos por Vallés et al. [29], quienes confirmaron que hay una selección in vitro de las células normales genéticamente para la formación de embriones somáticos, y que las plantas que se obtienen a partir de ellos son generalmente estables. Se ha reportado que la selección en el proceso de diferenciación desempeña un papel importante. Esto se determinó al comparar el número de cromosomas en las células del tejido de los callos y el nivel de ploidía de las plantas regeneradas, lo que pudiera ser la causa del desarrollo in vitro de plantas diploides a partir de callos que contienen células poliploides. Por esta razón, se plantea que solamente las células normales desde el punto de vista citológico retienen la capacidad de regeneración in vitro [30]. Para muchas especies del género Allium se ha reportado inestabilidad cariotípica de los callos cultivados, así como modificaciones en las plantas regeneradas a partir de ellos [17]. Sin embargo, la variación genética que se produce en los tejidos somáticos no se detecta fácilmente debido a la ausencia de marcadores fenotípicos adecuados en los cultivos de tejidos. La inestabilidad genética in vitro tiene grandes desventajas, entre ellas la incapacidad para regenerar un gran número de plantas idénticas (propagación clonal de genotipos), la disminución de la capacidad de regeneración, la regeneración de plantas estériles y la pérdida de marcadores genéticos [31], pero, a su vez, es de gran importancia cuando se quiere seleccionar nuevos individuos con características deseadas [2]. Estudio de la variación inducida por análisis de patrones de peroxidasas Por permitir el estudio directo del producto de los genes, el análisis isoenzimático constituye un método adicional para estudiar los cambios genéticos que ocurren durante el crecimiento y almacenamiento de los cultivos in vitro. Por esa razón, las diferencias en los patrones de bandas de isoenzimas entre los tejidos del cultivar original y los cultivados in vitro, pueden ser utilizadas para detectar tales cambios [32]. La Figura 5 muestra una representación esquemática de los patrones de isoperoxidasas correspondientes a los diferentes tejidos estudiados. Al analizar los patrones obtenidos, se observa, en general, que hay variación en el número e intensidad de las bandas para los callos y hojas de las plántulas regeneradas in vitro, en relación con el patrón obtenido para las hojas de las plantas del cultivar original. Figura 5. Patrones electroforéticos de las peroxidasas en los callos y las hojas de las plantas regeneradas in vitro. Para las hojas de las plántulas obtenidas a partir de semillas aparecen tres bandas solamente. De ellas, las bandas A2 y C1 aparecen en los demás tejidos estudiados (callos y plántulas regeneradas in vitro), aunque varían cuantitativamente en los diferentes tejidos y se observa la mayor intensidad en los callos. Por otro lado, la banda A1 aparece solamente en las plantas obtenidas a partir de semillas y su ausencia es general en las plantas regeneradas y los callos. Vale la pena destacar que los patrones obtenidos para los cultivos in vitro muestran muchas más bandas que el cultivar original; sin embargo, resulta interesante que sólo tienen en común las bandas que también están presentes en el cultivar original (A2 y C1). Las bandas A3, C2 y C4 sólo aparecen en las plantas regeneradas in vitro, mientras que las bandas C3 y C5 solamente están presentes en los tejidos de los callos. En los callos las bandas también son más intensas, lo que muestra una mayor cantidad de isoenzimas presentes en este tejido. Estos resultados coinciden con lo planteado por Sabir et al. [33], quienes señalan que el estudio de las proteínas y las enzimas para caracterizar la variación somaclonal se puede resumir en tres categorías: movilidad electroforética alterada, pérdida o ganancia de bandas, y niveles alterados de proteínas específicas (diferente intensidad en las bandas). La función de las isoenzimas peroxidasas ha sido muy discutida en la literatura por su relación con los mecanismos de resistencia a factores bióticos y/o abióticos [34], y también se ha utilizado ampliamente en estudios de variación somaclonal en callos y plantas regeneradas [35]. En la actualidad, existe gran variedad de métodos moleculares disponibles para caracterizar el genoma vegetal, y tienen, a su vez, un enorme valor en las investigaciones relacionadas con la detección de la variación somaclonal [36]. Entre estos trabajos, se pueden citar los estudios basados en diferentes técnicas como los marcadores RAPD [37], RFLP [25], los microsatélites [38] y AFLP [39]. La presencia de la variación genética inducida durante el cultivo in vitro, tiene una consideración especial en el contexto de la transformación genética. No obstante, esta situación se complica debido a que en plantas obtenidas mediante cultivo de tejidos hay tantos informes de estabilidad como de inestabilidad, de ahí la necesidad de realizar un estudio detallado en cada caso específico. Conclusiones
Referencias
Copyright Elfos Scientiae 2000 The following images related to this document are available:Photo images[ba00073b.jpg] [ba00073d.jpg] [ba00073e.jpg] [ba00073a.jpg] [ba00073c.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}