 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnología Aplicada, Vol. 18, No. 1, March 2001, pp. 1-9 Iron and Transferrin. Research and Therapeutic Applications
Antonio Aguila La O*
Department of Immunology and Bacteriology. University of Glasgow. Western , Infirmary.
Jeremy H. Brock
*Autor de correspondencia Received July 2000 Code Number: BA01001 Abstract Iron plays an important physiological role in eucaryotic and procaryotic cells, which have evolved complex mechanisms to ensure an adequate iron supply and tight regulation of its metabolism. In vertebrates a group of iron-binding proteins performs these functions, keeping concentrations of biochemically and biologically available iron far below the levels required for microbial growth and generation of cytotoxic free radicals. How these proteins make iron metabolism a broad and effective mechanism of antimicrobial defence and homeostatic balance is discussed. Here we review the microbial mechanisms for iron-uptake in the human bacterial pathogens Escherichia coli, Neisseria meningitidis, Haemophilus influenzae, and Staphylococcus aureus. We also address the implications for host-parasite relationships and the significance of iron as a regulatory signal for the expression of virulence factors. Iron also exerts an important influence on the function of the human immune system and on the development of neoplasia. Potential clinical, diagnostic and therapeutic applications of iron biochemistry are highlighted. In particular we suggest that serum transferrin, the major extracellular iron transporter in systemic fluids, should be considered a promising target for modulating iron metabolism in clinical conditions. Exploration of iron metabolism offers a new way of looking at microbial infections. Its advantages in clinical medicine, research of microbial pathogenesis, and drug development in the alarming emergence of bacterial antibiotic resistance have been just recognised. Keywords: iron, iron-binding proteins, lactoferrin, Neisseria meningitidis, neoplasia, transferrin, Staphylococcus aureus, vaccines. Resumen Hierro y transferrina. Aplicaciones en la investigación y la terapia. El hierro desempeña un papel importante en la fisiología de las células eucariotas y procariotas, las cuales han desarrollado mecanismos complejos para garantizar su suministro adecuado y control estricto. En los vertebrados, un grupo de proteínas fijadoras de hierro realiza estas funciones, manteniendo los niveles de hierro biológicamente disponibles muy por debajo de los niveles requeridos para el crecimiento microbiano y la generación de radicales libres citotóxicos. En el presente trabajo se analiza cómo estas proteínas hacen del metabolismo del hierro un mecanismo efectivo y universal de protección antimicrobiana, y de mantenimiento de la homeostasis. Los mecanismos microbianos de asimilación del hierro son descritos en patógenos humanos como Escherichia coli, Neisseria meningitidis, Haemophilus influenzae y Staphylococcus aureus, así como sus implicaciones prácticas en las interacciones hospedante-parásito y su significación en la regulación de la virulencia microbiana. El hierro también influye en el sistema inmune y en el desarrollo de neoplasias. Se resaltan las aplicaciones clinicas, diagnósticas, y terapeúticas de la bioquímica del hierro. La transferrina, el principal transportador de hierro extracelular en los fluidos sistémicos, debe ser considerada un objetivo importante en la modulación del metabolismo del hierro en condiciones clínicas. El estudio del metabolismo del hierro ofrece una nueva visión de las infecciones microbianas, cuyas ventajas en la clinica, la investigación de la patogenicidad microbiana, y el desarrollo de fármacos en el contexto de la alarmante emergencia de bacterias resistentes a antibióticos son finalmente reconocidas. Palabras claves: hierro, lactoferrina, Neisseria meningitidis, neoplasia, proteínas fijadoras de hierro, Staphylococcus aureus, transferrina, vacunas Note: Part of this review was presented in the Plenary Lecture "Iron and Microbial Growth", given by Dr. Brock on occasion of receiving the Membership of Honour of the Cuban Society for Immunology. Finlay Institute, Havana, July 14, 1999. Introduction Interest in the microbiostatic capacity of specific iron-binding proteins (IBP) against certain human pathogens declined 50 years ago [1], because of the growing success of recently discovered antibiotics. During this period the indiscriminate use of antibiotics in human and veterinary medicine has provoked a sharp and continued increase in bacterial antibiotic resistance [2]. Due to this alarming situation, the attention of the medical, research and pharmaceutical communities has turned once again to the study of the host-parasite interactions in an attempt to find alternatives to antibiotics. Attention has focused on the mechanisms of microbial virulence and their counterpart, the host. s mechanisms of natural resistance against microbial infection. As a result, a new picture has emerged, which points to iron and iron metabolism as one of the most important components of both microbial virulence and immune protection [3]. It is our aim to review iron metabolism from the point of view of host-parasite relationships, pointing out the mechanisms involved in the proper supply of iron for physiological needs. Iron metabolism is one of the most effective mechanisms of innate defence, as a reduction in biologically available iron prevents deleterious iron-catalysed oxidative reactions and microbial growth. We then focus on the bacterial mechanisms of iron uptake and discuss the vaccine potential of siderophore/transferrin receptor proteins in infections by important human bacterial pathogens. Finally, the implications of iron metabolism in infection and neoplasia are addressed briefly. The current and potential pharmaceutical applications of the research results in each of the topics mentioned above is discussed critically, and the most promising areas for future research are highlighted. Chemical Properties and Biological Functions of Iron in Eucaryotes and Procaryotes After the discovery of Schade and Caroline [1], a great deal of work focused on the biochemical activity of transferrins as iron transporters, and at first there was little interest in their antimicrobial effect. However, the discovery in the 1960. s of the capacity of iron compounds to enhance the virulence of pathogenic bacteria in vivo prompted a renewed interest in their antimicrobial effects [4]. This exclusive property of iron has been applied in animal models of bacterial infections [5, 6], the understanding of bacterial pathogenicity in vivo, and the unravelling of the clinical significance of alterations in iron metabolism on antimicrobial resistance mechanisms [3]. Iron is such a biologically important component in a large number of vital biochemical functions in procaryotic and eucaryotic organisms (Table 1) by two unique chemical properties: the reversible change between the ferrous (Fe2+) and ferric (Fe3+) oxidation states due to the unfilled electronic distribution of the external d atomic orbitals and the capacity to change its redox potential and structural arrangement according to the nature of the co-ordinating ligand. Table 1. Role of iron in microbial and mammalian biochemical processes
and examples
However, reactions of iron with oxygen can lead to production of harmful free radicals, highly reactive with cellular structures, DNA, etc. Consequently, aerobic organisms have evolved specific enzymes such as superoxide dismutase, gluthatione peroxidase, and catalase to counteract these negative effects (reviewed by Brock et al. in 7). In contrast, this potentially life-threatening property of iron can be of benefit during the "respiratory burst" in activated mammalian polymorphonuclear cells, where generation of free radicals helps killing of phagocytosed microorganisms [8]. Iron plays an important role in all living cells, which have evolved mechanisms to ensure an adequate supply of iron to meet their physiological needs. On one hand, the potential toxicity of iron, and its capacity to enhance the virulence of microbial pathogens, impose on eucaryotic organisms the need to restrict biochemically and biologically available iron to levels insufficient for both processes. On the other hand, procaryotic organisms, which require iron for multiplication and expression of their virulence factors during infection, must also develop effective mechanisms for iron uptake to overcome the limitations imposed by the host. The mechanisms of iron metabolism developed by vertebrates will be described in order to facilitate the understanding of their implications in microbial virulence, immunity and neoplasia. Iron Metabolism in Eukaryotes On a functional basis there are three main groups of IBP: (a) oxygen-binding proteins, (b) proteins catalysing redox reactions, and (c) iron-complexing proteins. The first group comprises proteins able to reversibly bind oxygen through a haem group, and consequently, transport oxygen between different cells. Haemoglobin is responsible for oxygen transport at cellular level whereas myoglobin stores it in muscle tissues [9]. Serum haemopexin and haptoglobin bind free haem groups and released haemoglobin respectively, and deliver them to the liver (haem groups) and kidneys (haemoglobin), where the complexes are eliminated by the reticuloendothelial system, thus preventing haem iron-catalysed generation of free radicals and limiting the availability of iron-loaded haem groups to pathogenic microorganisms. These acute phase proteins are involved in natural immune defence against microbial infections and tissue damage. The second group includes a very large number of enzymes which play a pivotal role in the metabolism of iron in all living organisms (Table 1). A detailed explanation of these proteins is beyond the scope of this review, and more comprehensive information may be found elsewhere [3]. Here we will simply point out that iron is central to the most essential and universal biological process: DNA replication, as it is the cofactor of ribonucleotide reductase, the enzyme responsible for reduction of all ribonucleoside diphosphates for DNA synthesis. In the third group, iron-complexing proteins regulate iron metabolism at intracellular and extracellular levels. In the intracellular milieu, ferritin, the major iron-storage protein in living cells, plays an important role in iron metabolism due to its ability to maintain large quantities of iron in a soluble, non-toxic and biologically available form. Ferritin is a hollow orange-shaped molecule comprising 24 subunits of about 20 kD each, which surround a core of up to 4500 ferric atoms. Mammalian cells produce two types of ferritin polypeptides distributed differentially in tissues. The H-subunit with a more acid pI predominates in non-iron storage organs, such as heart, red blood cells, lymphocytes and monocytes, while the L-subunit with a more basic pI predominates in iron storage organs, such as the liver, spleen and placenta [10]. This tissue distribution correlates with the suggested distinct roles of the H-rich and L-rich ferritin isoforms in iron detoxification and iron storage, respectively. The synthesis of ferritin is co-ordinated with the amount of extracellular iron and the expression of the transferrin receptor, a topic addressed later in this review. Although several mechanisms have been advanced, there is not yet a definitive explanation about how ferritin-bound iron is released intracellularly [7]. Extracellular ferric iron is bound by transferrins, a family of homologous monomeric two-sited iron binding glycoproteins of molecular weight about 80 kD with very high affinity for ferric iron (KD ~10-22 M) in co-ordination with bicarbonate anion at physiological pH values [3]. Transferrins can be classified according to their origin. Serum transferrins are found in all tissue fluids, whereas lactoferrins are present mainly in mucosal secretions and the granules of neutrophils. Ovotransferrins occur in egg whites, and a newer class, named melanotransferrin, was more recently found in the membranes of malignant melanoma cells. Transferrin, the most important protein involved in withholding and transportation of extracellular free iron, will be the focus of this review. The molecule is organised in two lobes with nearly 40% amino acid sequence homology between the N- and C-terminal lobes, which are thought to have arisen via gene duplication of an ancient single gene [11]. Each lobe is organised into two dissimilar subdomains separated by a cleft that contains the iron-binding site. The two-iron binding sites differ in their affinity for iron and their acid stability. The N-lobe loses iron at pH values between 5.5 and 6.0, whereas the C-lobe shows more acid stability, losing iron between pH 4.50 and 5.0. N-lobe iron-loaded and iron-free (apo) forms are the predominant variants of plasma transferrin. These properties may allow the fine tuning of iron release in different environments. In contrast to transferrin, lactoferrin activities are not related to iron transport, and although a detailed analysis of its properties is beyond the scope of this review, a comparison with transferrin is shown in Table 2. Table 2. Major differences between lactoferrin and transferrin.
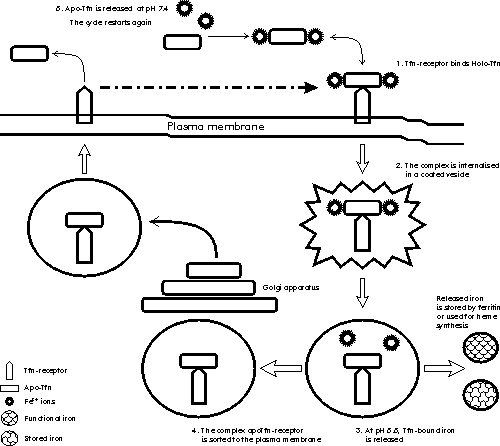
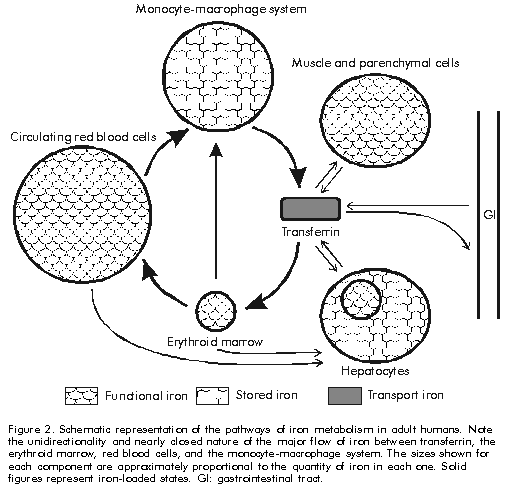
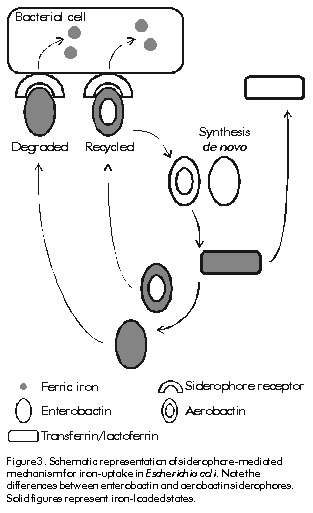
In the case of transferrin, a receptor-mediated endocytic pathway for iron release to cells has been described [7]. It involves the preferential internalisation of iron-saturated transferrin after binding to its cell surface receptor at the plasma membrane. This complex is taken up into vesicles by receptor-mediated endocytosis where acidification to pH 5.5 decreases the affinity between transferrin and iron, since binding of apotransferrin to its receptor is favoured at pH 5.0-6.5, whereas iron is released to the cell either for storage in ferritins or for synthesis in the mitochondrion. The transferrin-receptor complex is then sorted through Golgi apparatus to the plasma membrane. At extracellular pH, the transferrin receptor shows higher affinity for iron loaded transferrin, thus the complex dissociates and apotransferrin becomes available for recycling (Figure 1). Figure 1 . Schematic representation of the receptor-medited endocytic mechanisms for iron uptake from transferrin in vertebrates. See text for further details. Solid figures represent iron-loaded states. The concentrations of extracellular and intracellular iron are balanced by the coregulation of the expression of ferritin and transferrin receptor. A regulation mechanism based on the selective interaction of cytoplasmic iron-regulatory proteins to highly conserved 5' and 3' messenger RNA sequences of transferrin receptor and ferritin has been described [7]. Several less well understood mechanisms of iron regulation have been proposed in eucaryotes [3]. In healthy humans the total amount of iron is normally about 40 mg Fe/kg body weight in women and 50 mg Fe/kg in men. It is distributed in the following way: 65% in haemoglobin, 4% in myoglobin, 1% in the haem enzymes involved in intracellular oxidative processes, 0.1% in serum transferrin, and 15. 30% stored intracellularly in ferritins. Iron absorption of about 1 g of iron per day occurs in normal humans through duodenal epithelial cells and it is strictly related to body iron levels. There are not physiologically regulated mechanisms for the excretion of iron from humans. However, 0.05% of iron is lost daily in sweat, and in the ferritin within the gut epithelial cells that desquamate to the intestinal lumen, a factor of potential interest in the virulence of mucosal pathogens; menstruating women lose about 15. 20 mg monthly [12]. The cycle of iron metabolism is represented schematically in Figure 2. Figure 2. Schematic representation of the pathways of iron metabolism in adult humans. Note the unidirectionality and nearly closed nature of the major flow of iron between transferrin, the erythroid marrow, red blood cells, and the monocyte-macrophage system. The sizes shown for each component are approximately proportional to the quantity of iron in each one. Solid figures represent iron-loaded states. GI: gastrointestinal tract. Humans show the most effective system of iron conservation in any animal species, a phylogenetic advantage to be considered before extrapolating experimental results obtained in animal models to humans. Thus severe iron deprivation is one of the challenges that pathogenic microorganisms have to face to multiply and synthesise their virulence factors during infections in healthy animals. Microbial Mechanisms of Iron Uptake Iron uptake is a universal requirement for all microorganisms, pathogenic or not. Potential host sources of iron for invading pathogens are: IBP, ferritin, haemoglobin or haem, and intracellular pools. Here we will focus on the mechanisms for uptake of extracellular iron developed by pathogenic bacteria, where iron uptake has been much better characterised in Gram-negative than in Gram-positive species. Studies with protozoal and fungal pathogens are even scarcer and inconclusive [3]. In the extracellular milieu, partially iron-saturated IBP (from now onwards, the IBP acronyms will be used for referring only to transferrin and lactoferrin), are in equilibrium at concentrations as low as 10-18M free iron, a level insufficient to support microbial growth, and more severe hypoferremia is induced by innate mechanisms during infection, providing additional support to the role of iron deprivation in antimicrobial defence. It must be realised that it is not simply the lack of iron that microorganisms must confront during infections, but the biological unavailability of highly abundant iron, a situation not to be overlooked during in vitro and animal experimentation. In physiologically normal tissue fluids bacteria might be expected to assimilate ferric iron bound by IBP by the mechanisms proposed below. Transferrin is taken as model for IBP, while lactoferrin is mentioned where appropriate. Proteolytic cleavage of the IBP and release of the bound ferric iron have been described in some strains of Bacteroides fragilis and in bacteria associated with periodontiditis [13], but as only a few observations have been reported, it is not possible to conclude that it is a universal mechanism for iron uptake. Recently, interest has focused on the capacity of the Vibrio cholerae hemagglutinin/metalloenzyme (Hap) to degrade lactoferrin [14]. Clinical trials of genetically attenuated V. cholerae CTXf vaccine strains report adverse effects in human volunteers immunised orally with a CTXf, hap+ strain [15], but not with the CTXf, hap- strain [16], implying a role for Hap in cholera virulence. Experiments to elucidate how Hap-mediated proteolysis affects the antimicrobial activity of lactoferrin against V. cholerae are being carried out by our group [17]. As IBP have a much lower affinity for ferrous iron compared with ferric iron, the second mechanism proposes the reduction of the bound ferric iron and release of the ferrous form to pathogenic bacteria. Listeria monocytogenes and Escherichia coli have been shown to produce both soluble and surface-associated reductases which reduce and release ferric iron from transferrin and ferritin in vitro [18,19]. A similar activity was recently observed by our group in supernatants of Staphylococcus aureus cultured in chemically-defined medium as described in [17]. The reductase activity secreted by both bacteria was completely abolished by proteinase K treatment (1.5 mg/mL) suggesting its protein nature. Interestingly, transferrin-bound, but not lactoferrin-bound iron, was reduced when added to supernatants of E. coli and S. aureus, which correlates with the higher bacteriostatic effect of lactoferrin compared to transferrin (Aguila et al., manuscript in preparation). Although the physiological significance of this effect has not yet been proved, the presence of highly reducing conditions and low pH in bacterial abscesses [3] suggest it could occur in vivo. The third mechanism involves the generation of siderophores; low. molecular-weight iron-chelating organic compounds with extremely high affinity for ferric iron, whose secretion is induced under iron limiting conditions (Figure 3). E. coli secretes two types of siderophores of different chemical nature: enterochelin or enterobactin, and aerobactin [20]. Iron-loaded enterobactin is degraded completely after intracellular internalisation to release bound iron. In contrast, aerobactin is recycled, providing a comparatively less metabolically expensive pathway for iron uptake. There is a controversy about the functional advantages of the apparently redundant simultaneous production of two siderophores by the same bacterial strain. Brock et al. reported that advantages include different kinetics and effectiveness of iron uptake at different pH values, or iron concentrations, as well as differences in the iron source used by each siderophore, with enterobactin preferentially taking up IBP-bound iron, while aerobactin chelates iron from cells and tissues [21]. Figure 3. Schematic representation of siderophore-mediated mechanism for iron-uptake in Escherichia coli. Note the differences between enterobactin and aerobactin siderophores. Solid figures represent iron-loaded states. Siderophore-secreting bacteria (Table 3) also produce outer membrane protein receptors and a whole complex of transmembrane and cytoplasmic proteins that are involved in internalisation of iron-loaded siderophores [3, 20]. These intracellular proteins are not specific for any siderophore, which allows the cross utilisation of siderophores by all bacteria capable of expressing the specific siderophore receptor and provides the structural basis for development of siderophore-like molecules with targeted antimicrobial activities [22, 23]. Although specific antibodies against siderophore receptor proteins have been detected after bacterial infections, their protective value still awaits definitive evaluation. Table 3. Examples of bacterial pathogens and their mechanisms for iron-uptake.
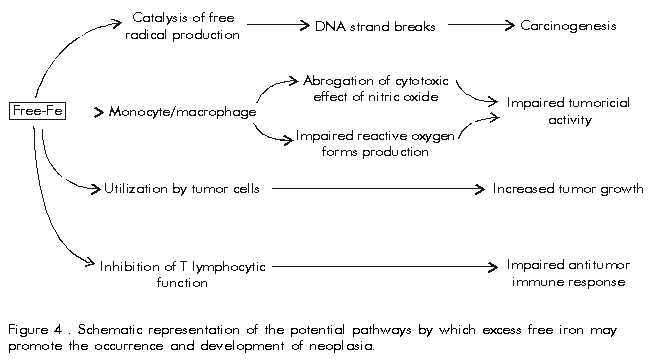
The fourth mechanism is based on direct contact between IBP and cognate bacterial cell surface receptors. Interestingly, despite the high amino acid sequence and structural homology between IBP, there are individual, specific bacterial receptors for each of them, and in contrast to siderophores, the IBP-receptor. based mechanisms are highly species-specific for the IBP of a particular host [3, 24]. We will describe this mechanism in the case of major human pathogens. Generally, host-restricted bacteria as Neisseria meningitidis and Haemophilus influenzae express only IBP-receptors, while biphasic (i.e., both environmental and pathogenic) bacteria such as S. aureus produce siderophores and perhaps IBP-receptors as well (Table 3). N. meningitidis expresses two transferrin binding proteins, TbpA (Tbp1) and TbpB (Tbp2) of 98 and 70 kD, respectively. While TbpB can be renatured after isolation and detected by immunoblotting; TbpA cannot be, and demonstration of an interaction between transferrin and this Tbp requires previous binding in situ to intact cell membranes using appropriately labelled transferrins [24]. A detailed explanation of the mechanism of iron uptake from transferrin by meningogoccal Tbps is beyond the scope of this review; and comprehensive information may be found elsewhere [25]. There seem to be crucial differences in the mechanism of iron acquisition from lactoferrin, as although two lactoferrin-binding proteins LbpA (Lbp1) and LbpB (Lbp2) have been identified in Neisseriacea, LbpB is not essential for uptake of lactoferrin-bound iron in vitro [26]. Further studies are needed to elucidate the physiological significance of this observation. H. influenzae is a major cause of bacterial meningitis, second only to N. meningitidis. Transferrin-binding proteins are induced in H. influenzae cultured in iron-deprived media supplied with protoporphyrin IX [27]. The transferrin receptors of H. influenzae also consist of the Tbp proteins. In contrast, lactoferrin-binding proteins seem to be uncommon in these bacteria. Two proteins isolated by lactoferrin affinity chromatography lacked the conserved sequences typical of Tbp or Lbp leading to the conclusion that they are not functional lactoferrin-receptors. This seems a paradox, given the mucosal tropism of this pathogen. In contrast to N. meningitidis and H. influenzae, S. aureus is a ubiquitous microorganism capable of causing a variety of human and veterinary diseases. The wide host range and tropism of S. aureus may be related to the presence of both siderophore-mediated [28] and transferrin receptor-mediated [29] mechanisms for iron uptake. A single 42 kD Tbp was recently identified, which happens to be the only known bacterial Tbp with glyceraldehyde-3-phosphate dehydrogenase (GAPDH) enzymatic activity. This unusual activity in bacterial Tbp suggests a potential contribution of GAPDH activity to bacterial virulence far beyond iron acquisition, but this question still awaits definitive elucidation [29]. In contrast to Gram-negative Tbps, staphylococcal Tbp is able to chelate iron from single-lobed transferrin fragments, preferably from the N-lobe which makes iron particularly accessible to blood-borne staphylococci. Interestingly, S. aureus gapdh- strains, lacking GAPDH activity, showed a reduced capacity to use holotransferrin as a source of iron, which may suggest either the presence of a second staphylococcal transferrin receptor or of separate domains for enzymatic activity and transferrin binding (P. Williams, unpublished observations). Although the presence of S. aureus lactoferrin-binding proteins has been reported [30], the absence of further information about their cellular location, role in iron-uptake, and level of regulation by iron, means it is not yet possible to consider them as functional lactoferrin receptors. Expression of Tbps is always iron-regulated in N. meningitidis, but H. influenzae [27] and S. aureus [29] strains show constitutive Tbp expression. The functional significance of this difference in genetic regulation of Tbp expression may lie in the ability of invasive bacteria to constantly face an iron-deprived environment in tissue fluids. These results have been applied to try and develop vaccines against N. meningitidis serogroup B and H. influenzae type b [5, 6]. Several groups have focused their efforts on the potential use of transferrin-binding proteins as vaccine antigens against N. meningitidis. There are several reports that Tbps are immunogenic in humans after natural infections or asymptomatic carriage [31], as well as in animals immunised with purified Tbps [32]. Animals immunised with Tbp preparations produced bactericidal and opsonic anti-Tbp IgG antibodies, as well as antibodies capable of blocking iron uptake by N. meningitidis, and were effectively protected against lethal challenge with several virulent homologous and heterologous meningococcal strains [33]. The usefulness of Tbps as meningococcal vaccine antigens has been hampered by their considerable antigenic and genetic heterogeneity, which may lead to antigenically complicated, and therefore expensive, vaccine formulations. Genetic analysis of Tbp and the observation that in contrast to animal models, human anti-Tbp antibodies show a considerable degree of crossreactivity and crossprotection, suggest that transferrin receptors can be grouped into two major families [34, 35]. It seems that the inclusion of correctly folded Tbp from each meningococcal Tbp group will increase both the level of protection and the spectrum of strains against which protection can be achieved in a single vaccine formulation. However, it should not be overlooked that vaccine formulations containing other outer membrane proteins have induced effective protection in mass vaccination programmes in humans, which has been critically analysed by Tappero et al. [36]. The vaccine potential of bacterial lactoferrin receptors has not been subjected to such intensive research as their transferrin counterparts. However, studies with N. meningitidis [37] have demonstrated the immunogenicity of Lbp2 in vivo, and support their potential as vaccine antigens. However, as with Tbps, their genetic and antigenic heterogeneity must be assessed before considering the inclusion of Lbp in a meningococcal vaccine formulation [38]. Iron also regulates the expression of bacterial virulence factors, which might be unrelated to iron metabolism, such as toxins, haemolysins, adhesive and motile factors, etc. Iron-dependent global gene regulators. the repressor protein Fur (ferric uptake regulation), and tRNA-modification. functioning in co-ordination with other environmental signals: temperature, osmotic strength, pH, oxygen levels, and general nutrient availability to enable invading pathogens have been described, and their implication in the environmental monitoring by invading pathogens, discussed [3, 7]. There are still complete fields with promising therapeutic and prophylactic applications, such as the mechanisms of iron acquisition by intracellular pathogens, the interactions between bacterial mechanisms of virulence and host immune defences, and their implications in bacterial pathogenesis, the potential of bacterial IBP/siderophore receptors for development of vaccines and specifically targeted-drugs, the role of iron as a global regulatory signal of the expression of virulence factors. Changes in iron metabolism may have decisive effects on the final outcome of host-pathogen interactions. IBP-mediated suppression/retardation of microbial growth is essential for microbial killing by the innate and adaptive immune defences. The high incidence of microbial infections in hyperferremic and severely traumatised subjects as well as the natural induction of body hipoferremia during infections validate this frequently overlooked fact. Neglect of this mechanism has caused unnecessary problems, as the infection-promoting effects of the haemoglobin substitutes developed without considering the iron-binding properties of natural haemoglobin [3, 7]. A recent publication linking siderophores, IBP and phagocytes as a mechanism of host defence seems to indicate a positive move towards this direction [39]. Iron in Immunity and Neoplasia Specific immunity involves a series of metabolically demanding processes starting from antigen recognition and lymphocyte activation and proliferation to cytokine-mediated immune regulation and cellular co-operation [8]. The metabolic requirement of iron for all such processes (Table 1) and the defective immune responses observed in humans and experimental animals with abnormalities of iron metabolism, extend the implications of iron metabolism from innate to adaptive immunity. Although clinical studies in humans have been hindered by difficulties, such as lack of appropriate controls, and influence of concurrent diseases and physiological abnormalities, a general pattern has emerged about the role of iron in immunity and neoplasia. Impairment of T-cell functions shown by reduced cutaneous sensitivity to appropriate model antigens in humans, and defects in T-cell response to mitogens in animals have been associated with iron deficiency. The low iron saturation of serum transferrin found in such conditions may result in an inadequate supply of iron for the activation and proliferation of T-cells. This explanation is supported by the observation that primed T-cells from iron deficient donors are capable of mounting a proper immune response after adoptive transfer to iron normal recipients [7]. T-cell subsets seem to differ in their dependence upon iron, with a greater inhibitory effect of iron deficiency on Th1 than Th2 clones. This suggests that changes in iron status could influence the induction and regulation of immune responses [40]. Similar studies with B lymphocytes are less convincing, as contradictory effects on B-cell activation and normal antibody production have been reported in patients with different levels of iron. Overall, B-cell activation seems to be less iron-dependent that T-cell activation. In both lymphocytic populations transferrin-bound iron has a positive impact on proliferation reflecting a common requirement for iron. In contrast, free iron and non-transferrin bound iron forms inhibit lymphocyte proliferation in vitro. This effect has been explained by an uncontrolled uptake of iron by lymphocytes, which are unable to synthesise enough ferritin for iron storage, leading to membrane damages and DNA strand breakage caused by free radicals generated by outer membrane and/or intracellularly located iron polymers. On the other hand, Brock et al. showed that the cytotoxic activity of macrophages and lymphocytes is reduced during iron deficiency due to the role of iron in the generation of both oxygen- and nitrogen-reactive intermediaries [41]. That iron overload may predispose to neoplasia is supported by the increased incidence of hepatocellular carcinoma in patients with the severe form of hereditary haemochromatosis, as well as the significant association between iron accumulation in the liver and the development of hepatic cirrhosis in patients with chronic active Hepatitis C [42]. Tumor cells can overgrow normal cells by two main distinctive properties: the use of non-transferrin bound extracellular iron, overcoming its cytotoxic effects in hyperferremic environments such as in chronically Hepatitis C virus infected liver, and the synthesis of high quantities of transferrin, which favours their growth in the iron-deprived and poorly vascularized areas surrounding actively growing tumors. Taken together, these observations suggest that free iron may not only initiate the carcinogenic process, but may also favour the selective growth of tumor cells over other cells such as those of the immune system consequently impairing their antitumoral effects [7] (Figure 4). These results have found practical application in the design of specifically targeted drug delivery systems based on the specificity and enhanced expression of transferrin receptors in tumor cells. Enhanced allograft survival has been shown in heart-transplanted mice treated with antitransferrin receptor and anti IL-2 receptor antibodies [43], while new possibilities for the treatment and/or diagnosis of neoplasia based on the specific delivery of labelled, and biologically active compounds, such as suicide genes, hammerhead ribozymes, liposomes, interferons, etc. coupled to antitransferrin receptor antibodies have been addressed [44]. Figure 4. Schematic representation of the potential pathways by which excess free iron may promote the occurrence and development of neoplasia. Conclusions Iron plays an important physiological role in eucaryotic and procaryotic cells, which have evolved complex mechanisms to ensure an adequate iron supply and tight regulation of its metabolism. In higher organisms, a whole set of IBP carries out such functions, while keeping concentrations of biochemically and biologically available iron far below the levels required for generation of cytotoxic by-products, and microbial growth. Microbial pathogens have been forced to develop specific mechanisms to confront this nutritional limitation, making iron and iron metabolism a broad and effective mechanism of antimicrobial defence and of homeostatic balance, as well as an important regulatory signal in the expression of virulence factors in vivo. There are fields of research, such as the rigorous clinical assessment of the effect of iron status on the immune system of vertebrates, microbial virulence, siderophore/IBPs bacterial receptors, development of neoplasias, etc, which would repay future studies by generating diagnostic, clinical, therapeutic, and prophylactic applications in immune regulation, microbial pathogenesis, vaccine development, cancer treatment, etc. In particular we suggest that serum transferrin, the major extracellular iron transporter in systemic fluids, should be considered a promising target for modulating iron metabolism in clinical conditions. This review aims to provide a new way of looking at microbial infections, where the appropriate consideration of iron metabolism as a component of the innate and adaptive immunity is still more important in view of both the alarming results of the antibiotic era: increasing antibiotic resistance, and incompleted understanding of the mechanisms of host resistance and microbial virulence; and of the recently discovered and promising pharmaceutical applications based on iron metabolism. Acknowledgements Support by the Royal Society, United Kingdom, the Cuban Society for Immunology and the Finlay Institute is gratefully acknowledged. We thank Mrs. Francesca Marcantonio, Royal Society, London and Dr. Gustavo Sierra, Finlay Institute, Havana, for their kind encouragement and helpful assistance. Our gratitude to Prof. Allan M. Mowat, Department of Immunology and Bacteriology, University of Glasgow for critical reading of the manuscript. References
Copyright Elfos Scientiae 2001
The following images related to this document are available:Line drawing images[ba01001b.gif] [ba01001c.gif] [ba01001a.gif] [ba01001d.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}