 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnología Aplicada, Vol. 18, No. 1, March 2001, pp. 10-16 Aplicaciones de la calorimetría diferencial de barrido al estudio de la estabilidad de las proteínas Alejandro Beldarraín
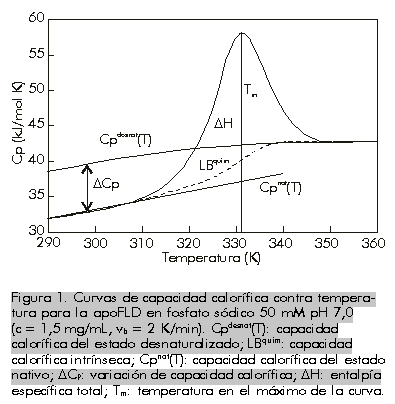
Recibido en agosto del 2000. Aprobado en noviembr del 2000. Code Number: BA01002 RESUMEN En este manuscrito, se analizan las posibilidades de aplicación de la calorimetría diferencial de barrido al estudio de la estabilidad de las proteínas. Se describe la metodología de la técnica cuando los procesos de desnaturalización térmica son de equilibrio, como aquellos casos en los que existe un control cinético. Se ofrecen también las ecuaciones necesarias para el tratamiento de los datos, aplicadas a algunas proteínas producidas en la Planta de Producción del Centro de Ingeniería Genética y Biotecnología de La Habana, Cuba. Palabras claves: apoFLD, desnaturalización térmica, estabilidad de proteínas, rHu-gIFN, rSk ABSTRACT Application of Differential Scanning Calorimetry to Protein Stability Studies. In this manuscript, the possibilities of application of the differential scanning calorimetry to the study of protein stability are analyzed. The methodology of the technique is described when the processes of thermal denaturation are in equilibrium, like the cases in which a kinetic control exists, and the necessary equations for data treatment are given, applied to some proteins produced in the Production Plant of the Center for Genetic Engineering and Biotechnology in Havana, Cuba. Keywords: apoFLD, protein stability, rHu-gIFN, rSk, thermal denaturation Introducción Una de las principales prioridades de la industria biotecnológica moderna es la obtención de moléculas estables o estabilizadas; es decir, moléculas resistentes a los efectos de la temperatura, a los agentes caotrópicos, a los solventes orgánicos, a los detergentes, etc. A esto se adicionan las estrategias de estabilización de estructuras moleculares para prevenir su inactivación biológica por cualquier agente externo, asunto al que se le dedican esfuerzos considerables en la actualidad. Estos aspectos están relacionados directamente con la falta de comprensión del mecanismo de plegamiento de las biomoléculas in vivo; o sea, la falta de conocimientos sobre la segunda parte del código genético. La organización compleja y la estabilidad de la estructura tridimensional de una biomolécula, se pueden investigar experimentalmente in vitro a través del estudio del proceso de su destrucción; es decir, del estudio de su desnaturalización y de los factores externos que influyen. El método calorimétrico es el único que no necesita suponer a priori los mecanismos de estos procesos. La técnica de calorimetría de barrido es la más eficiente para su estudio, debido a que proporciona información muy certera acerca de los parámetros termodinámicos de estabilización de la estructura macromolecular [1]. En los últimos años, la calorimetría diferencial de barrido (CDB) de alta sensibilidad se ha convertido en una herramienta poderosa para caracterizar los cambios conformacionales inducidos por la temperatura en las macromoléculas biológicas, y se ha empleado fundamentalmente en estudios de predicción del mecanismo de plegamiento de las proteínas. Esta técnica ha servido para determinar con exactitud los factores que influyen en la estabilidad de las biomoléculas [1-11]. En los últimos años, se han publicado trabajos sobre la aplicación de la CDB en análisis cuantitativos y semicuantitativos de la estabilidad de compuestos farmacéuticos [6, 12, 13], y en la evaluación de aditivos de formulaciones líquidas de compuestos de gran valor terapéutico [14, 15]. El objetivo de este trabajo fue hacer una explicación de esta técnica con la inclusión de ejemplos que demuestren su potencialidad a partir del estudio de proteínas recombinantes, entre ellas algunas producidas por el Centro de Ingeniería Genética y Biotecnología de La Habana, Cuba. ¿Qué es la calorimetría diferencial de barrido? Esta técnica es relativamente sencilla. Mediante la CDB se mide la capacidad calorífica relativa de un sistema determinado cuando sufre una transición inducida por un cambio térmico [1, 3, 4]. En el experimento, las celdas con la muestra y la referencia se calientan simultáneamente a una misma velocidad predeterminada. La muestra es una solución o una suspensión del material que se desea estudiar, y la referencia contiene solamente el disolvente utilizado. Cuando se calientan ambas celdas y se produce una transición inducida por la temperatura (por ejemplo, la desnaturalización de una proteína o una transición orden-desorden en una bicapa fosfolipídica), la muestra absorbe parte del calor que se le está suministrando a la celda. Como resultado, la célula de la muestra tendría una temperatura algo inferior a la de la celda de referencia, pero el sistema de control del equipo suministra una potencia calorífica adicional para que ambas celdas mantengan la misma temperatura. Esta potencia en exceso es proporcional a la diferencia de capacidades caloríficas entre las dos celdas y sus contenidos. De esta forma simple se obtiene la capacidad calorífica del soluto en función de la temperatura. En la Figura 1 se muestra un experimento típico de CDB aplicado a una proteína. Se puede observar que el valor de capacidad calorífica de la proteína Cp, no es el mismo antes y después del proceso de desnaturalización, con un valor de DCp > 0. Esta capacidad calorífica intrínseca, LBquim, se resta de la curva de capacidad calorífica anterior para obtener la capacidad calorífica de exceso, Cpexc (Figura 2). La reversibilidad de la transición se comprueba normalmente con el recalentamiento de la muestra de proteína en la celda calorimétrica, inmediatamente después de enfriar la celda que se calentó en el primer barrido. Esta prueba de reversibilidad sólo muestra que el sistema vuelve al estado inicial (nativo) después del enfriamiento, pero es posible que el sistema no esté en equilibrio todavía si está controlado cinéticamente por la velocidad de barrido utilizada. Una prueba de reversibilidad mejor consiste en comprobar que la curva obtenida no depende de la velocidad con que se varía la temperatura [16]. Tanto para esta comprobación como para la realización de cualquier experimento de CDB, conviene siempre asegurar que la respuesta del instrumento sea lo suficientemente rápida para que no se distorsione la señal (o al menos que se pueda corregir por los métodos de corrección dinámica existentes) [7]. En la referencia 17 se puede encontrar una descripción detallada de un microcalorímetro diferencial de barrido y de la metodología experimental que se debe seguir para su utilización. Figura 1. Curvas de capacidad calorífica contra temperatura para la apoFLD en fosfato sódico 50 mM pH 7,0 (c = 1,5 mg/mL, vb = 2 K/min). Cpdesnat(T): capacidad calorífica del estado desnaturalizado; LBquim: capacidad calorífica intrínseca; Cpnat(T): capacidad calorífica del estado nativo; DCP: variación de capacidad calorífica; DH: entalpía específica total; Tm: temperatura en el máximo de la curva. Figura 2. Curva de capacidad calorífica de exceso para la apoFLD fosfato s ódico en 50 mM pH 7,0. A la curva experimental (línea continua) se le restó la línea base, la línea química y está ajustada por mínimos cuadrados no lineales a una curva teórica de dos estados (línea discontinua). Procesos de equilibrio La información que se puede obtener de un experimento de CDB es diferente cuando el proceso está en equilibrio durante el barrido de temperatura (transición reversible), en comparación con un proceso controlado cinéticamente (transición irreversible). En los casos de reversibilidad en las desnaturalizaciones seguidas por CDB, se acepta, por lo general, la aplicación de la termodinámica de equilibrio para el análisis de los datos. Modelo de equilibrio de dos estados La desnaturalización más simple es aquella en la que solamente los estados nativo (N) y desnaturalizado (D) están poblados significativamente, y sus cantidades relativas a una temperatura determinada están determinadas por el valor de la constante de equilibrio de desnaturalización K:
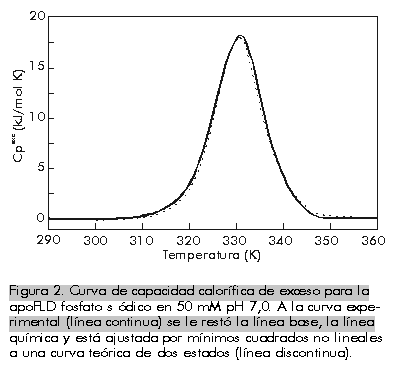
donde: A este modelo se ajustan proteínas pequeñas comúnmente [1]. La Figura 2 es una representación de una curva de capacidad calorífica de exceso obtenida para la apoFLD, a pH neutro y a baja fuerza iónica. Esta curva fue calculada para un proceso de equilibrio simple entre dos estados cuando el sistema sufre un barrido de temperaturas en el rango en que ocurre esta transición, a través de la ecuación: suponiendo que DCp = 0, Tm = 330,45 K y DH = 263 kJ/mol. La entalpía del proceso en estudio (DH) es simplemente el área bajo la curva, que a veces se conoce como entalpía total de la transición o entalpía calorimétrica (DHcal), pues: donde: La temperatura en el máximo de la curva (Tm) coincide prácticamente con la temperatura a la que se produce la transición justo a la mitad, con poblaciones similares de los estados nativo y desnaturalizado. Para este proceso, el calor absorbido a una temperatura determinada es proporcional al avance de la reacción. Por lo tanto, la constante de equilibrio del proceso (K) a cada temperatura se puede obtener fácilmente a partir de esta curva, y a partir de la ecuación de van't Hoff se puede calcular el cambio de entalpía (DHvH) correspondiente del proceso N D, según: Se puede demostrar de forma simple que a T = Tm [5]:
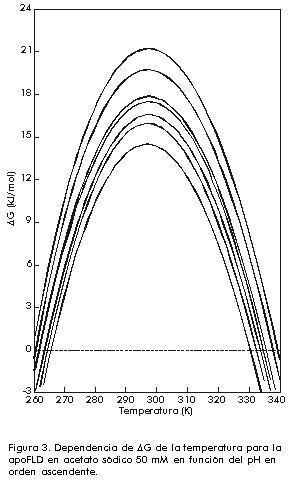
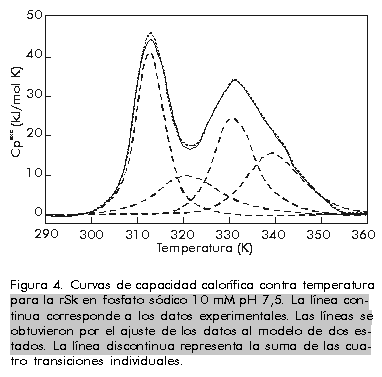
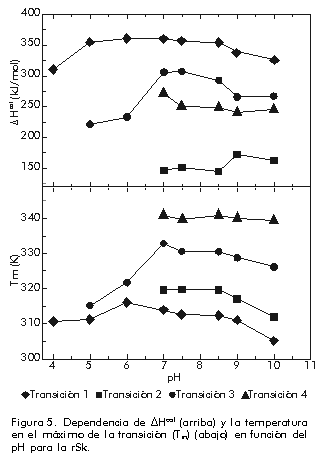
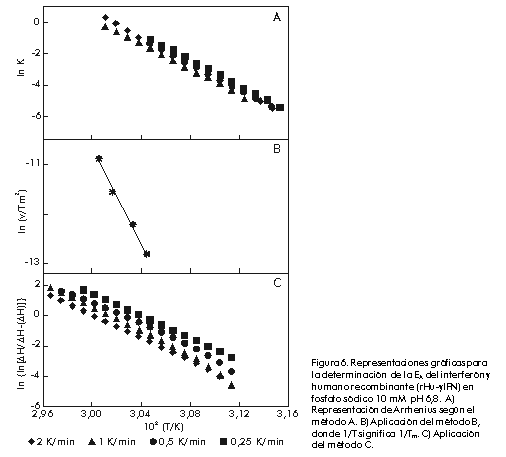
donde: Si este proceso es, como se ha supuesto, un equilibrio entre dos estados A y B, los valores de DHvH y DHcal deben ser idénticos. De hecho, DHvH carece de sentido si éste no es el caso. Sin embargo, la razón r = DHcal/DHvH, que debe ser igual a 1 en este modelo, se utiliza muy a menudo para comprobar si el modelo se cumple o no, e incluso para obtener información del proceso que se está estudiando (vea más adelante). Una vez que se conocen los valores de DCp y DH a T = Tm, a partir de un experimento único, es posible obtener los cambios de entalpía (DH), entropía (DS) y energía de Gibbs (DG) a otras temperaturas para el proceso entre los dos estados, a partir de ecuaciones termodinámicas conocidas [1]: DH(T) = DH(Tm) + DCp(T-Tm) En la Figura 3 se muestra una representación de DG(T) para la apo-FLD en función de la temperatura como medida de la estabilidad conformacional y en función del pH. Se observa que los valores de la energía de Gibbs son bajos (entre 20 y 80 kJ/mol) los cuales representan una compensación entre valores de entalpía y entropía de un orden de magnitud superior. A cada temperatura, los valores de DG se pueden interpretar como una medida de la estabilidad de la proteína en estudio, si se admite que todas las proteínas tienen, aproximadamente, un estado desnaturalizado de igual energía como se comprobó hace ya bastantes años [18]. También se puede observar en esta figura que la estabilidad (o DG) es nula a Tm y que, además de una desnaturalización por calor, se puede producir también una desnaturalización por frío, hecho que se ha comprobado de forma independiente para otras proteínas [4]. Figura 3. Dependencia de DG de la temperatura para la apoFLD en acetato sódico 50 mM en función del pH en orden ascendente. Existe cooperatividad intermolecular cuando se observa que DHvH > DHcal, aunque el proceso siga siendo de equilibrio. Un caso extremo es el de una transición de primer orden, en el que r tiende al infinito. Cuando una proteína es un dímero en ambos estados, el nativo y el desnaturalizado, r tiende a 2. Si la proteína es un n-ámero en el estado nativo, pero está totalmente disociada en el estado desnaturalizado, se puede demostrar que a T = Tm: r = 4[(n-n1/2)/(n-1)]2 En el caso de las transiciones térmicas conocidas de las bicapas fosfolipídicas, la razón r indica el número medio de moléculas de lípido que sufren la transición, a las que se le llama tamaño de la unidad cooperativa [19]. Además del análisis termodinámico explicado anteriormente, se puede obtener una medida de la estabilidad conformacional de cualquier biomolécula en función de las condiciones del medio, si se toman como referencia la temperatura en el máximo de la transición (Tm) y la entalpía calorimétrica que caracteriza al proceso de desnaturalización. Equilibrio entre más de dos estados El hecho de que DHvH<DHcal en el proceso de equilibrio, indica que este proceso no es de dos estados y que existen intermedios estables termodinámicamente [2]. Esta situación corresponde normalmente al caso en que ciertas regiones de la proteína, o dominios cooperativos, se desnaturalizan más o menos independientemente uno del otro. Se han propuesto varios modelos para explicar estos casos. El más antiguo y más frecuentemente usado es el modelo secuencial, que consiste en varios estados consecutivos durante el proceso de desnaturalización [20]. Los parámetros termodinámicos de cada estado se pueden calcular a partir de la curva experimental Cp(T) por ajuste de mínimos cuadrados no lineales, por el análisis en etapas recursivas en función de la temperatura hasta que se obtengan las constantes de equilibrio, la fracción de moléculas en cada estado, los cambios de entalpía y cualquiera otra función relacionada con ellos. En la Figura 4 se muestra un ejemplo de un sistema analizado por este método, correspondiente a la transición térmica de la estreptoquinasa recombinante (rSk) a pH neutro. Figura 4. Curvas de capacidad calorífica contra temperatura para la rSk en fosfato sódico 10 mM pH 7,5. La línea continua corresponde a los datos experimentales. Las líneas se obtuvieron por el ajuste de los datos al modelo de dos estados. La línea discontinua representa la suma de las cuatro transiciones individuales. La deconvolución del termograma (líneas discontinuas en la Figura 4) muestra que la proteína tiene cuatro dominios que se desnaturalizan independientemente en función del pH con diferente estabilidad (Figura 5). Aunque los dominios sean termodinámicamente estables e independientes, la estabilidad global de la proteína y, por consiguiente, su actividad biológica, puede estar determinada por la estabilidad del dominio más sensible. Por lo tanto, se hace indispensable conocer cuál es la parte de la estructura que corresponde a este dominio. Una manera bastante efectiva que se ha utilizado, incluso para la estreptoquinasa, es la fragmentación por digestión proteolítica y la evaluación posterior del fragmento obtenido, con la comparación de su comportamiento calorimétrico con el observado para la proteína íntegra [21, 22]. Figura 5. Dependencia de DHcal (arriba) y la temperatura en el máximo de la transición (Tm) (abajo) en función del pH para la rSk. Otro aspecto interesante que vale la pena destacar es la cantidad de dominios de esta proteína, que varía de 1 a 4 según algunos reportes [21-26]. La posibilidad de que pequeños cambios en las condiciones experimentales o en la estructura de la misma proteína puedan influir en la cantidad de dominios, ha sido analizada por Beldarraín y colaboradores [27]. Otro modelo al cual se ajustan muchos datos experimentales bastante bien es el llamado modelo de transiciones independientes [28]. Este modelo supone que la desnaturalización de una proteína es un proceso de desnaturalización de cada uno de sus dominios y que estos procesos son independientes unos de otros; es decir, que la desnaturalización de un dominio no influye en la desnaturalización de los demás. En los últimos años, se han introducido otros modelos [8, 29] que tienen en cuenta el hecho de que los diversos dominios interaccionan más o menos entre ellos. Esto se suele hacer con la introducción de una energía de Gibbs de interacción (DGij) entre los dominios i y j. Este modelo coincide con el modelo de transiciones independientes cuando DGij es muy grande. Procesos de no equilibrio Los equilibrios descritos anteriormente no se cumplen para muchas proteínas solubles y para ninguna de las proteínas de membrana estudiadas hasta la fecha. Por lo tanto, los parámetros termodinámicos que se obtengan mediante la aplicación de la metodología descrita anteriormente pueden ser falsos, excepto el área debajo de la curva que representará todavía la entalpía de desnaturalización del proceso, pues las curvas calorimétricas están altamente distorsionadas por el efecto de la alteración irreversible. La desnaturalización térmica irreversible es atribuida generalmente a alteraciones del estado final, como la autólisis, las agregaciones, las alteraciones químicas de los residuos de aminoácidos, etc., que impiden que la proteína adopte su estructura nativa. Estas alteraciones irreversibles son fundamentalmente procesos cinéticos que se pueden describir a través de ecuaciones de velocidad. Las curvas que se obtienen normalmente en casos de no equilibrio son similares a las que se muestran en la Figura 6 (líneas continuas). Sin embargo, también en este caso es posible obtener información útil acerca del proceso de desnaturalización, aunque será, claro está, de carácter cinético y no termodinámico. Para ello se pueden aplicar diversos modelos cinéticos que permiten el análisis de las curvas experimentales de CDB [7, 16, 30]. Aquí se describe simplemente el más sencillo de todos: el modelo de Lumry y Eyring [16]. Figura 6. Representaciones gráficas para la determinación de la EA del interferón g humano recombinante (rHu-gIFN) en fosfato sódico 10 mM pH 6,8. A) Representación de Arrhenius según el método A. B) Aplicación del método B, donde 1/T significa 1/Tm. C) Aplicación del método C.
donde: El equilibrio entre los estados N y D se describe mediante la constante de equilibrio K y la etapa irreversible es de primer orden con una constante de velocidad kF. Este modelo de Sánchez-Ruiz (N®F) permite determinar la constante de velocidad (kF), y su energía de activación (EA), relacionadas por la ecuación de Arrhenius:
donde: El modelo conduce a una serie de ecuaciones mediante las cuales se puede calcular EA: A) La constante de velocidad de la reacción, a una temperatura dada, se puede calcular mediante la ecuación:
donde: Mediante el uso de la ecuación de Arrhenius, se puede representar ln k contra a 1/T para los valores de k obtenidos a cada temperatura. Todos los valores deben quedar en una recta independientemente de la velocidad de barrido a la que se han obtenido. En la Figura 6A se muestra la representación para el interferón g recombinante (rIFN-g ). B) El modelo predice que la temperatura del máximo (Tm) debe cambiar con la velocidad de barrido; según la ecuación:
de manera que, si se representa ln(v/Tm2) contra 1/Tm, se debe obtener una recta de pendiente -EA/R. Esta representación para el rIFN-g se muestra en la Figura 6B. C) El calor desarrollado a cada temperatura <DH> es: por lo que se puede obtener EA de una representación del primer término de esta ecuación contra 1/T, que debe ser una línea recta de pendiente -EA/R. En la Figura 6C se muestra esta representación para el rIFN-g. D) También se puede calcular EA directamente a partir de la ecuación:
donde: Cuando estos cuatro métodos conducen a valores coincidentes para la energía de activación, se puede concluir que se cumple el modelo. Sin embargo, es preferible comprobar los resultados experimentales con los valores calculados (ver el modelo E). E) Un quinto método se deduce a partir de la derivación de la ecuación del modelo C y teniendo en cuenta la ecuación del modelo D:
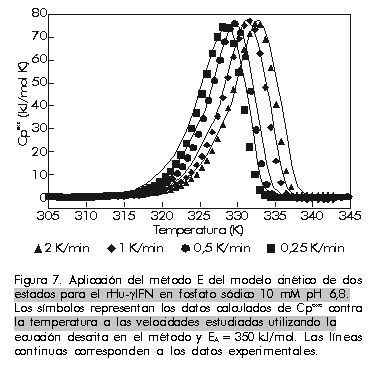
La energía de activación se puede obtener mediante un ajuste no lineal de mínimos cuadrados de los datos experimentales de Cp (T) (Figura 7). Figura 7. Aplicación del método E del modelo cinético de dos estados para el rHu-gIFN en fosfato sódico 10 mM pH 6,8. Los símbolos representan los datos calculados de Cpexc contra la temperatura a las velocidades estudiadas utilizando la ecuación descrita en el método y EA = 350 kJ/mol. Las líneas continuas corresponden a los datos experimentales. Se ha visto que las transiciones dependen en gran medida de la velocidad de barrido en este modelo, como se había mencionado antes para procesos de no equilibrio. Ésta es una regla general independientemente del modelo que se emplee para explicar los datos de CDB de desnaturalizaciones controladas cinéticamente. Por lo tanto, se puede emplear como criterio de aplicación o no de la termodinámica de equilibrio cuando se encuentren transiciones irreversibles desde el punto de vista calorimétrico. Este modelo tan sencillo ha descrito muy bien los datos de CDB de proteínas que se desnaturalizan de forma irreversible [31-35]. En la Tabla y la Figura 7 se puede apreciar la buena concordancia entre los valores experimentales y las curvas teóricas calculadas para el rIFN-g . Tabla. Valores de energía de activación obtenidos por diferentes métodos para
la desnaturalización térmica
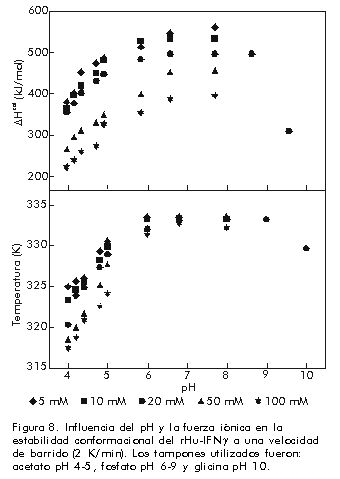
Es obvio que se puede extraer información cinética sólo de la aplicación de este modelo, ya que la información termodinámica queda totalmente enmascarada por la presencia del proceso irreversible. No obstante, para una velocidad de barrido específica, el valor de Tm se puede utilizar como parámetro comparativo para evaluar la estabilidad de una biomolécula en función de las condiciones del medio (Figura 8). Figura 8. Influencia del pH y la fuerza iónica en la estabilidad conformacional del rHu-IFNg a una velocidad de barrido (2 K/min). Los tampones utilizados fueron: acetato pH 4-5, fosfato pH 6-9 y glicina pH 10. Conclusión La CDB puede proporcionar información valiosa acerca de la naturaleza del desplegamiento de una macromolécula y de las fuerzas que influyen en la estabilidad del estado nativo. Por esa razón esta técnica se puede emplear ampliamente para la evaluación rápida y efectiva de moléculas nuevas o modificadas con relevancia biotecnológica, producidas por técnicas de ingeniería genética . ya sea para uso clínico o industrial. y constituye una herramienta indispensable en el momento de elegir una estrategia para la obtención de una molécula muy estable térmicamente en condiciones biológicas óptimas. Aunque la utilidad de esta técnica es indudable, es muy conveniente utilizarla junto a otras técnicas calorimétricas, en particular la calorimetría de titulación isotérmica o con la calorimetría de flujo, pues estos métodos pueden proporcionar información adicional en el examen de, por ejemplo, la termodinámica de las interacciones entre macromoléculas y ligandos específicos en función de la temperatura, en estudios de oligomerización de proteínas o, simplemente, en la evaluación de la influencia de la estabilidad de una macromolécula en una formulación específica. Agradecimientos Este trabajo ha sido realizado con la ayuda del Ministerio Español de Ciencias gracias a una beca otorgada a AB (Acción Especial BIO95-2068-E) en la Unidad de Resonancia Magnética Nuclear del Instituto Pluridisciplinar de la UCM. También agradecemos la colaboración de las Divisiones de Estreptoquinasa e Interferones de la Planta de Producción del CIGB que suministraron parte de las proteínas estudiadas. Referencias
Copyright Elfos Scientiae 2001
The following images related to this document are available:Line drawing images[ba01002f.gif] [ba01002t.gif] [ba01002h.gif] [ba01002m.gif] [ba01002j.gif] [ba01002k.gif] [ba01002d.gif] [ba01002e.gif] [ba01002c.gif] [ba01002n.gif] [ba01002l.gif] [ba01002p.gif] [ba01002i.gif] [ba01002q.gif] [ba01002o.gif] [ba01002r.gif] [ba01002a.gif] [ba01002s.gif] [ba01002g.gif] [ba01002b.gif] | ||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
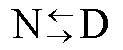{kind=link}
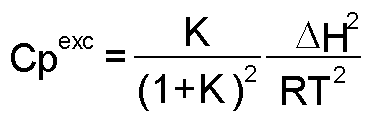{kind=link}
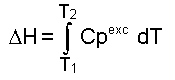{kind=link}
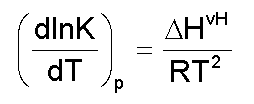{kind=link}
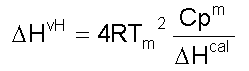{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
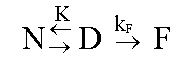{kind=link}
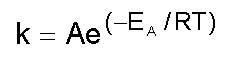{kind=link}
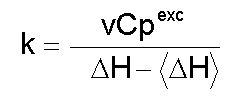{kind=link}
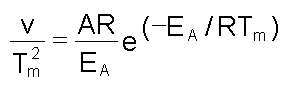{kind=link}
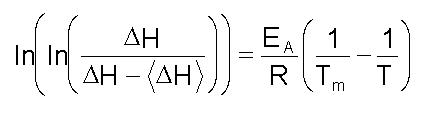{kind=link}
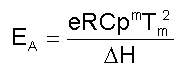{kind=link}
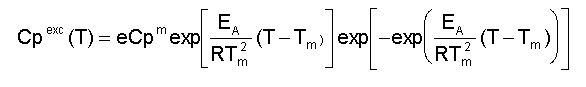{kind=link}
{kind=link}
{kind=link}