 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
SHORT ORIGINAL PAPER ARTICULO RORIGINAL CORTO INFLUENCE OF AAV-2 TERMINAL REPEATS ON EXPRESSION OF HETEROLOGOUS PROMOTERS. Julio Fernandez^1, Hilke Muller^2, Michael Ruffing^2 and Jurgen A. Kleinschmidt^2. ^1 Centro de Ingenieria Genetica y Biotecnologia. Apartado postal 6162, Habana 6. Ciudad de La Habana, Cuba. ^2DKFZ, ATV, Heidelberg, Germany.
Recibido en septiembre de 1994. Aprobado en febrero de 1995. Key words: AAV-2, heterologous promoters, terminal repeats. SUMMARY
The adeno-associated virus is a human parvovirus that can be used as a vector system for gene transduction into mammalian cells. For integration into the host cell genome, replication and packaging of the viral DNA only terminal repeats are necessary in cis. We investigated the influence of the terminal repeats on the expression of genes under the control of heterologous promoters in transient transfection assays. The activity of HPV-18, HCMV and HIV-1 promoters in vectors with and without terminal repeats was compared using luciferase as a reporter gene. We observed a reduction of gene expression from all promoters tested in plasmids with terminal repeats. The inhibitory effect was abolished with the expression of one viral regulatory protein and infection with adenovirus 5. RESUMEN
El virus adeno-asociado es un parvovirus humano que puede ser utilizado como vector para la transduccion genica en celulas de mamiferos. Para la integracion en la celula hospedera, la replicacion y el empaquetamiento del ADN viral, solo se requieren de las secuencias terminales repetidas en cis. Nosotros estudiamos la influencia de estas secuencias en la expresion de genes bajo el control de promotores heterologos en experimentos de transfecion transiente. La actividad de promotores de HPV-18, HCMV y HIV-1 en vectores con y sin terminales repetidos, se comparo usando la luciferasa como gen reportero. Se observo una disminucion de los niveles de expresion con todos los promotores usados en plasmidios que contienen la secuencia de los terminales repetidos. El efecto inhibitorio se elimino con la expresion de una de las proteinas reguladoras del virus y la infeccion con adenovirus 5. INTRODUCTION The adeno-associated virus type 2 (AAV-2) is a human parvovirus that requires a helper virus (adeno or herpes virus) to replicate efficiently (1). The AAV-2 has a linear single stranded DNA genome of 4.6 kb with inverted terminal repeats (TRs) at each end. The TRs form a secondary structure that function as origin of replication. The TRs of about 125 bp are the unique sequences required in cis for replication, rescue from plasmids, packaging of AAV-2 DNA into virions and proviral integration (2-7). In the absence of helper virus, wild type AAV-2 integrates via TRs in a site-specific manner into human chromosome 19q (8). The left half of the AAV-2 genome contains the rep open reading frame, which encodes at least four overlapping regulatory proteins (Rep78, Rep68, Rep52, and Rep40). The four Rep proteins share a common reading frame and differ at their N and C termini depending on promoter usage and splicing. Rep proteins are required for efficient AAV-2 DNA replication and the production of single-stranded progeny for encapsidation (9-11). The right half of the AAV-2 genome contains the open reading frame for capsid proteins VP1, VP2 and VP3. They are expressed from overlapping sequences of the same open reading frame by using alternative initiation codons (12). The AAV-2 has been suggested as a vector for gene therapy protocols (13,14). The advantages for this are the following: First, the AAV-2 vector has been reported as a vector with high transduction frequencies in cells of diverse lineages, including hemopoietic cells (15,16). Second, the AAV-2 vectors have a simple structure compared with other viral vectors. Third, since the AAV-2 vector is not lytic and not replicating; antigens may be processed and presented optimally by an intact cell with small perturbation of normal cellular processes. Fourth, because integration occurs; its provide stable gene expression. In this paper we described the influence of the AAV-2 TRs in the expression of luciferase as a reporter gene under the regulation of heterologous promoters in transient transfection assays. MATERIALS AND METHODS Construction of recombinant plasmids
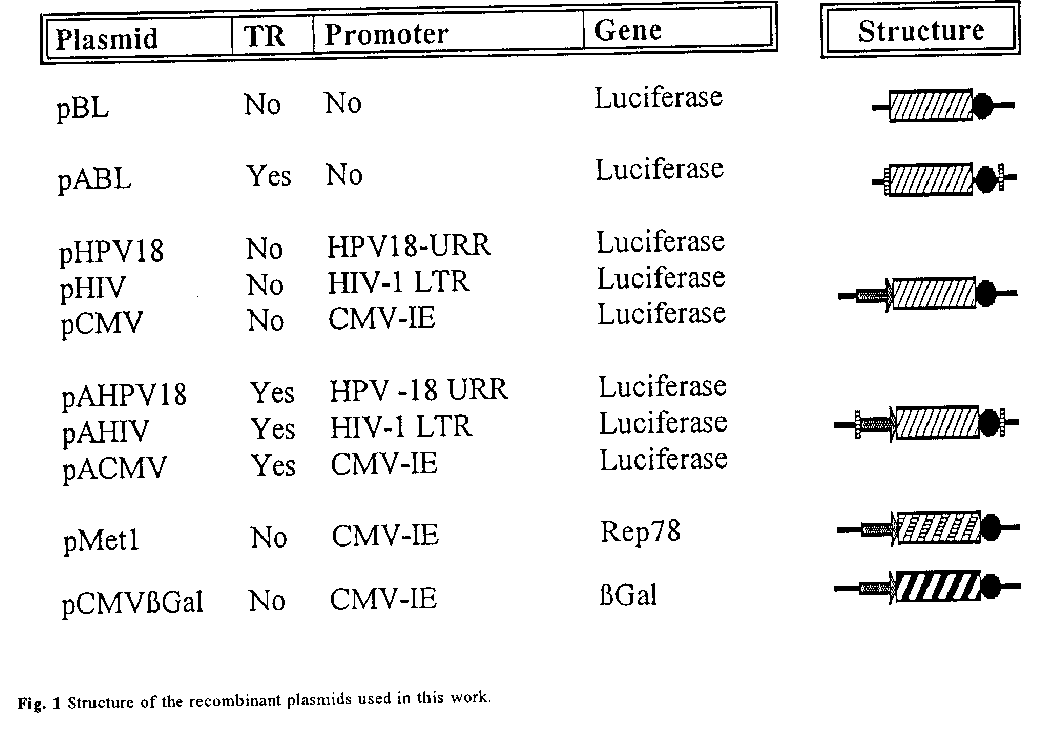
The plasmid pBL contains the luciferase gene from P.pyralis and the SV40-polyA in the Hind III-Xho I sites of the Bluescript plasmid (Stratagene, USA). This plasmid was used as a vector for cloning different viral promoters using the standard techniques (17). For cloning was used the 825 bp viral fragment from HPV-18 that contains the upstream regulatory region of the E6-E7 promoter (18), a 650 bp fragment from the HCMV corresponding to the enhancer-promoter region of the major immediate early gene (19), and a 1 100 bp fragment from HIV-1 that include the LTR regulatory elements (20). Each promoter and the luciferase gene SV40-polyA were cloned in the plasmid pSSV9 that contains the AAV-2 TRs (21). The plasmid pABL was constructed by deletion of the HPV-18 upstream regulatory region. Plasmid pMet1 contains the Rep78 gene under the control of the HCMV promoter, and pCMVbetaGAL contains the betaGal gene under the control of the HCMV promoter (22). They were kindly supplied by Markus H”rer. The plasmids are represented in figure 1. Large scale preparations of plasmids were obtained by growing the transformed E.coli strain HB101 in LBA broth. Plasmids were then purified using Quiagen columns as described by the manufacturers.
Cell culture and virus stock
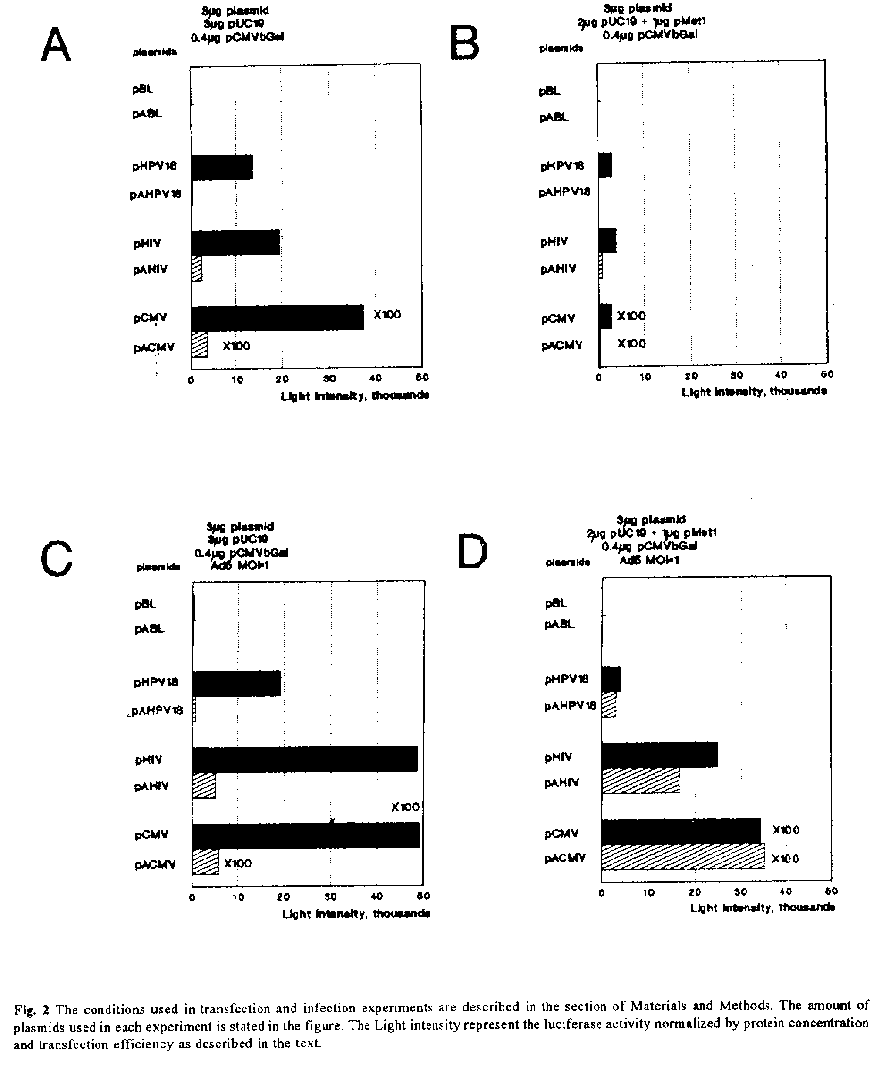
HeLa cells were propagated in Dulbeccos modified Eagles medium with 5% fetal calf serum, 100 mg each of penicillin and streptomycin per mL and 2 mM glutamine at 37^oC and 5% CO2. Adenovirus 5 (MOI=1) was used in experiments for infection of HeLa cells (21).
Transfection of HeLa cells Transfection of HeLa cells was performed by the protocol of Chen and Okayama (23). The day before transfection, 5 X 10^5 HeLa cells were seeded in a petri dish (diameter 6 cm) with 3 mL of medium and incubated at 37øC and 5% CO2. Mixtures of recombinant DNA (6.4 ug per dish as specified in individual experiments) was added to 150 uL of 2 X BBS buffer (23) and 150 uL of 250 mM CaCl2. After 20 min incubation at room temperature the transfection solution was added to the cells, and the cells were incubated for 18 h at 35øC and 3% CO2. After the supernatant had been removed, cells were washed twice with serum-free medium and then incubated in fresh medium with fetal calf serum for 24 h at 37øC and 5% CO2. In the experiments in which infection was carried out the adenovirus 5 (MOI=1) was added then. Luciferase assay, protein determination and beta-Gal assay Luciferase activity in transfected cells was determined as described before (24). Cell extracts were made at 24 h after transfection in 300 uL lysis buffer (1% Triton X 100, 25 mM Glycylglycin, 15 mM MgSO4, 4 mM EGTA, 1 mM DTT, 10% Glycerin, pH 7.8). Cell extracts (50-100 uL) were added to 100 mL of luciferase reaction buffer (25 mM Glycylglycin, 15 mM MgSO4, 4 mM EGTA, pH 7.8) and the luciferase activity was measured by the addition of 100 uL of luciferase buffer (0.152 mM Luciferin, 25 mM Glycylglycin, 15 mM MgSO4, 4 mM EGTA, 8 mM ATP, pH 7.8) in a Luminometer (Lumat LB 9501, Berthold). The protein concentrations of cell extracts were measured at 595 nm (ELISA-Photometer, Titerkek Multiscan Plus) by the method of Bradford (25) using a Bio-Rad protein assay kit. The beta-Gal activity was measured by the addition of 5-50 uL of cell extracts to 200 mL of reaction buffer (0.1 M NaHPO4, 1.1 mM MgCl2, 50 mM beta-Mercaptoethanol, 0.01% ONPG (w/v)) at 405 nm (ELISA-Photometer, Titerkek Multiscan Plus). RESULTS AND DISCUSSION As a very convenient internal control to measure transfection efficiency, we cotransfected the cells with a beta-Gal expressing vector (pCMVbetaGAL), and measured beta-Gal activity using the same lysate. Promoter activities were determined by calculating the luciferase/beta-Gal ratios. The conditions and results from several experiments, using at least two different plasmid preparations, are summarized in figure 2a-2d. To eliminate the possibility that other elements in the plasmid could influence in the luciferase activity, we measured the luciferase activity of plasmids pABL and pBL in all the experiments. As shown in figure 1, these plasmids contain all the elements except the promoter. Both constructions displayed basal levels of luciferase activity (figure 2a-2d), comparable with the background of HeLa cells when the pUC19 was used for transfection (data not shown). This activity remains unalterable in presence of Rep78 and adenovirus 5 infection. These results are in agreement with the observation that the TRs appear to be devoid of transcriptional regulatory elements (26). Recently, Flotte et al. (27) shown that the TRs sequence can function as a promoter for expression of a reporter gene in a transient assay or for the expression of cystic fibrosis transmembrane regulator in a long term stable integration assay. However, the interaction of transcription factors with the TRs is not documented (28). The TRs are expected to do not affect the expression of foreign genes under the control of heterologous promoters in the absence of rep genes and adenovirus (26). Therefore, vectors without rep genes (rep^-) have been used to drive the expression of foreign genes. These include the SV40 early promoter (21), the HCMV promoter, the murine sarcoma virus LTRs and the HIV-1 LTRs (29). We examined the transient expression of luciferase gene under the control of different viral enhancer/promoter elements when the AAV-2 TRs is present or not in cis. We shown in figure 2a, that the presence of TRs in cis have a negative effect in the level of expression of all promoters tested in HeLa cells. The possibility that the TRs affect the binding of cellular factors involved in transcription is not supported by in vitro experiments described elsewhere (28). These experiments shown a DNA-protein complex specific for the AAV-2 TRs only in AAV-2 infected extracts, and when the TRs adopt a covalently closed or hairpin configuration. Further experiments to study the mechanism of this inhibition should be done. The control of AAV-2 gene expression is complex; both helper virus and AAV-2 genes are involved. Therefore, we decided to study the possible influence of the Rep78 protein and adenovirus proteins on the expression levels when recombinant AAV-2 is used. In contrast to rep^- vectors, the behaviour of foreign control elements in rep^+ vectors is less predictable. Without adenovirus infection, the inhibitory effects on gene expression of rep gene products from a variety of viral and cellular promoters, including those of AAV-2 itself, have been reported in rep^+ vectors (30-32). To study the influence of an AAV-2 regulatory protein, we cotransfected the cells with the plasmid pMet1, which expressed only Rep78 (unpublished results). We observed a reduction of the luciferase expression in vectors with TRs about two-sixfold, but more dramatically reduction (three-tenfold) of the luciferase activity was observed when the TRs were not present (figure 2b). As we shown here, the inhibition by rep gene product affects in more degree vectors that do not contain the TRs. According to these results, the negative effect of Rep78 on promoters is reduced when the TRs is present in cis. In which form Rep78 might affect the gene expression is not clear. Directly binding of Rep proteins to viral promoters has not been described. One explanation is the possible binding of Rep78 with factors that bound to promoter regions. The reduction of the negative effect observed in these experiments could be explained by the binding of Rep78 to the TRs. According to this, the TRs could compete for binding of Rep78 with these factors reducing the negative effect of Rep78 on promoters, or the binding of Rep 78 to TRs reduces the inhibition produced by the presence of the TRs. It is possible that both process occurs simultaneously. The adenovirus proteins are important for the expression of AAV-2 proteins (26). To study the possible effect of these proteins on the AAV-2 TRs we used the adenovirus 5 for infection of HeLa cells in cotransfection experiments. The infection with adenovirus 5 resulted in the increase of the luciferase activity 1.5-2 times (figure 2c) with respect to the control experiment without infection (figure 2a). The differences of activities were maintained between plasmids with and without TRs, for this reason we believed that the increase in activities is due to an increase in the transfection efficiencies and is not related with the interaction of adenoviral proteins on promoters. This observation coincides with the use of adenoviruses to increase the efficiency of gene transfer into a broad spectrum of eukaryotic cells (33). Our results shown that the TRs do not contain sequences regulated by adenovirus proteins. We shown in the figure 2d a result of infection of HeLa cells with adenovirus 5 previously cotransfected in the presence of pMet1. We observed a reduction of the luciferase activity about two-five times in plasmids without TRs compared with conditions in figure 2c, but the activities remain in the same range when compared with conditions in figure 2a. The negative regulation of Trs in heterologous promoters was reverted in the presence of Rep78 and adenovirus infection. These results might reflect replication and amplification of the TRs containing plasmids or reversion of the negative regulation of TRs in heterologous promoters by the presence of Rep78 and adenovirus infection. We concluded that the level of expression of AAV-2 vectors is negatively regulated by the presence of Trs and this effect is overcome by the presence of Rep 78 and adenoviruses infection. This pattern of regulation by Rep and adenoviruses with heterologous promoters is observed also for the own AAV-2 promoters (26). The Trs should contain regulatory sequences that might interfere with internally placed promoter or enhanced elements. This negative effect of Trs on different promoters should be considered when AAV-2 is used as a vector for gene transfer experiments. The evidence of the same pattern of regulation in other cell lines is under study. REFERENCES. 1. ATCHINSON, R.W., B.C. CASTO, and W.M. HAMMON (1965) Science 149: 754-756. 2. JANIK, J.E., M.M. HUSTON, and J.A.ROSe (1981) Proc. Natl. Acad. Sci. U.S.A. 78: 1925-1929. 3. HAUSWIRTH, W.W., and K.I. BERNS (1977) Virology 79: 488-499. 4. LUSBY, E.W., R. BOHENZKY, and K.I. BERNS (1981) J.Virol. 37: 1083-1086. 5. SAMULSKI R.J., K.I. BERNS, M. TAN, and N. MUZYCZKA (1982) Proc. Natl. Acad. Sci. U.S.A. 79: 2077-2081. 6. SAMULSKI, R.J., L.S. CHANG, and T. SHENK (1987) J.Virol. 61: 3096-3101. 7. SAMULSKI, R.J., A. SRIVASTAVA, K.I. BERNS, and N. MUZYCZKA (1983) Cell 33: 135-143. 8. SAMULSKI, R.J. (1993) Curr-Opin-Genet-Dev. 3: 74-80. 9. SRIVASTAVA, A., E.W. LUSBY, and K.I. BERNS (1983) J.Virol. 45: 555-564. 10.HERMONAT, P., M.A. LABOW, R. WRIGHT, K.I. BERNS, and N. MUZYCZKA (1984) J.Virol. 51: 329-339. 11. CHEJANOVSKY, N., and B.J. CARTER (1989) Virology 173: 120-128. 12. BECERRA, S.P., J.A. ROSE, M. HARDY, B. BAROUDY, and C.W. ANDERSON (1985) Proc. Natl. Acad. Sci. U.S.A. 82: 7919-7923. 13. MULLIGAN, R.C. (1993) Science 260: 926-932. 14. FLOTTE T.R, (1993) J-Bioenerg-Biomembr. 25: 37- 42 15. TRATSCHIN, J.D., I.L. MILLER, M.G. SMITH, and B.J. CARTER (1985) Mol. Cell. Biol. 5: 3251-3260. 16.MURO-CACHO, C.A., R.J. SAMULSKI, and D. KAPLAN (1992) J.Immunotherapy 11: 231-237. 17. SAMBROOK, J., E.F. FRITSCH, and T. MANIATIS (1989) Molecular cloning: a laboratory manual. 2^nd Edition. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. 18.GIUS, D.,S. GROSSMAN, M.A. BEDELL, and L.A. LAIMINS (1988) J.Virol. 62: 665-672. 19.BOSHART, M., F. WEBER, G. JAHN, K. DORSCH-HASLER, B. FLECKENSTEIN, and W. SCHAFFNER (1985) Cell 41: 521- 530. 20. RATNER, L., W. HASELTINE, R. PATARCA, K.J. LIVAK, B. STARCICH, S.F. JOSEPHS, E.R. DORAN, J.A. RAFALSKI, E.A. WHITEHORN, K. BAUMEISTER, L. IVANOFF, S.R. PETTEWAY, M.L. PEARSON, J.A. LAUTENBERGER, T.S. PAPAS, J. GHRAYEB, N.T. CHANG, R.C. GALLO and F. WONG-STAAL (1985) Nature 313: 277-313. 21. SAMULSKI, R.J., L.S. CHANG, AND T. SHENK (1989) J.Virol. 63: 3822-3828. 22. MCGREGOR, G.R, and C.T. CASKEY (1989) Nucleic Acid Res. 17: 2365-2370. 23. CHEN, C., and H. OKAYAMA (1987) Mol. Cell. Biol. 7: 2745-2752. 24. BRASIER, A.R., J.E. TATE, and J.F.HABENER (1989) Biotechniques 7: 1116-1122. 25. BRADFORD, M.M (1976) Anal. Biochem. 72: 248- 254. 26. MUZYCZKA, N. (1992) Currents Topics in Microbiology and Immunology. 158: 97-129. 27. FLOTTE, T.R, S.A. AFIONE, R. SOLOW, M.L. DRUMM, D. MARKAKIS, W.B. GUGGINO, P.L. ZEITLIN and B.J. CARTER (1993) J. Biol. Chem. 268: 3781-3790. 28.DONG-SOO and N. MUZYCZKA (1989) J.Virol. 63: 3095-3104. 29. LEBKOWSKI, J.S, M.M, MCNALLY, T.B, OKARMA and L.B, LERCH (1988) Mol. Cell. Biol. 8: 3988-3996. 30. CHATTERJEE, S, P.R, JOHNSON and K.K, WONG (1992) Science. 258: 1485-1458. 31. LABOW M.A, L.H, GRAF, K.I, BERNS (1987) Mol. Cell. Biol. 7: 1320-1325. 32. ANTONI B.A, A.B, RABSON, I.L, MILLER, J.P, TREMPE, N, CHEJANOVSKY, B.J, CARTER (1991) J.Virol. 65: 396- 404. 33. HERZ J, and R.D. GERARD (1993) Proc. Natl. Acad. Sci. USA. 90: 2812-2816. Copyright 1995 Sociedad Iberolatinoamericana de Biotecnologia Aplicada a la Salud
The following images related to this document are available:Line drawing images[ba95009b.gif] [ba95009a.gif] |
| |||||||||

{kind=link}
{kind=link}