 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
ARTICULO ORIGINAL CORTO / SHORT ORIGINAL PAPER HIGH LEVEL EXPRESSION OF HUMAN IFN-alpha2b IN Pichia Pastoris Julia N. Garcia, Jorge A. Aguiar, Miriela Gill, Armando Alvarez, Juan Morales, Joel Ferrero, Beatriz Gonzalez, Gabriel Padron and Alfredo Menendez. Center for Genetic Engineering and Biotechnology, P.O. Box 6162, La Habana 6, Cuba Recibido en octubre de 1994. Aprobado en julio de 1995
Key Words: IFN-alpha2b, Pichia Pastoris, AOX1 promoter, acetylation
SUMMARY The gene coding for human interferon-alpha2b was integrated into the Pichia pastoris genome. The expression of the interferon gene was controlled by methanol-regulated alcohol oxidase (AOX1) promoter. An expression level of 400 mg of interferon per liter of culture was achieved under appropriate fermentation conditions. The molecule was purified to homogeneity and partially characterized in terms of biological activity and structural properties. Fast Atom Bombardment Mass Spectrometry (FAB-MS) allowed the verification of the aminoacid sequence and indicated the presence of amino-terminal acetyl groups in 70% of the final product. An alternative to eliminate this modification is suggested.
RESUMEN El gen que codifica para el interferon-alfa 2b humano fue integrado en el genoma de Pichia pastoris. La expresion del gen de interferon fue controlada por la induccion con metanol del promotor alcohol oxidasa 1. Se alcanzaron 400 mg de interferon por litro de cultivo en fermentacion. La molecula fue purificada y parcialmente caracterizada. Se verifico la secuencia aminoacidica por espectrometria de masas con ionizacion por bombardeo con atomos acelerados (FAB-MS) detectandose un 70% de acetilacion. Una alternativa para eliminar esta modificacion es propuesta.
INTRODUCTION Interferons (IFNs) are a family of pro>teins which are classified basically in three antigenically distinct groups, designated IFN-alpha, IFN-beta and IFN-ganma (1). They have shown to exert a multitude of biological effects such as antiviral, antiproliferative and immunoregulatory action (1). At least 15 functional genes in the human genome have been identified to code for proteins of the IFN-alpha family, which include IFN-alpha2. It is a molecule of 165 amino acids, containing four cysteine residues involved in the formation of two disulphide bridges (2). The gene coding for IFN-alpha2 has been cloned by several laboratories and expressed to moderately high levels using different host-vector systems such as Saccharomyces cerevisiae, Bacillus subtilis, Streptomyces lividans and Escherichia coli (3). The methylotrophic yeast P. pastoris has been described as a host for high-level heterologous gene expression upon genetic transformation with plasmids carrying those genes under the control of the methanol-regulated alcohol oxidase (AOX1) promoter (4). In a defined minimal medium containing methanol, P. pastoris can be grown to high cell density, reaching concentrations of 130 g of dry weight of cells per liter of fermentation broth (4). That regulation of gene expression by methanol was shown to be simple, relatively easy to scale up, and cost-effective for industrial purposes (4). In this paper we describe the integration of the human interferon-alpha2b gene into the P. pastoris genome, the efficient expression of this gene by the recombinant yeast strain, the purification and a preliminary characterization of the molecule.
MATERIALS AND METHODS Strains and media E. coli MC1066 strain was used for plasmid manipulation and amplification. The yeast strain P. pastoris MP36 HIS3- (5) was used as the host for the expression of IFN. LB (6), G0 (7), and YP (yeast extract 1g/L, peptone 20 g/L) media were supplemented with 50 ug/mL ampicillin, 2% glycerol, and 2% glucose, respectively. Vector construction and transformation Recombinant DNA techniques were performed according to Sambrook et al. (6). The oligonucleotides were synthesized according to Jimenez ewal. (8). Figure 1 summarizes the construction of the expression vector, pIFNPP. The IFN gene was obtained by PCR (9) from the pDA8 plasmid (3), using the oligonucleotides: 5'TGTGATCTGCCTCAAACCCACAGCCTG3' and 5'TCATTCC-TTACTTCTTAAACTTTCTTG3'. The 495 bp coding fragment was phosphorilated and cloned into pNAO407 previously digested with NcoI-EcoRI and treated with the Klenow fragment of E. coli DNA polimerase I. P. pastoris MP36 HIS3- was transformed with ClaI-SalI digested pIFNPP according to Hinnen et al. (10). Culture conditions Transformed yeasts were grown up to a cell density of 5 x 10^7 cells/mL of culture (OD600 1.0) in 5 mL of G0 media, supplemented with 2% glycerol. The cells were then washed with sterile water and transfered to 100 mL of the YP medium supplemented each 24 hours with 0.5% methanol as the sole carbon and energy source. The growth was achieved at 30 C with constant vigorous shaking, during 120 h. Fermentations were carried out on 5 liter fermentors (B. E. Marubishi), with 3 liters of effective volume, containing G0 medium supplemented with 2% glycerol. The initial optical density was 0.3 OD600 and the fermentation conditions were 700 rpm, 1vvm of aeration, 30 C and pH 5.0. Interferon expression was induced after 12 h of culture by addition of methanol and maintaining its concentration at about 0.4% during 120 h. Purification of biologically active IFN-alpha2 Induced cells were harvested by centrifugation at 3 500 g for 30 min. All steps were performed at 4 C. 80 g of cells were resuspended in 300 mL of lysis buffer (Tris HCl 0.1M pH 7.9, sucrose 10%, NaCl 0.3M, EDTA 5 mM). Cells were broken by mechanical shaking (Dyno Mill Model KDL, retention time 5 min) and the debris harvested by centrifugation at 3 500 g for 30 min. The pellet of broken cells (1.3 g of total protein) was solubilized with 90 mL of 30 mM Tris HCl, 30 mM NaCl, 5 mM DTT pH 7.2 and 6M GuCl at 4 C during 2 hours. The insoluble material was removed by centrifugation at 8 000 g for 30 min. IFN-alpha2 was recovered by slow dilution of the 6M GuCl solution with 1 200 mL of cold 30 mM Tris HCl, 30 mM NaCl, 80 uM CuSO4 pH 7.0 over the course of 6 h. IFN-alpha2 was purified from the diluted Guanidium chloride solution using a column of immobilized monoclonal antibody CB-IFN A 2.4 (11). The protein peak eluted from this column was concentrated 10 times by ultrafiltration (AMICON). Total protein concentration was determined by the Coomassie blue method (12). SDS-PAGE and Western blot were done according to Laemmli (13) and Towbin (14), respectively. The concentration of IFN in all samples was determined by ELISA (15). Interferon-induced antiviral activity of the final preparations was determined through the inhibition of the cytopathic effect of Mengovirus on Hep-2 cells (16).
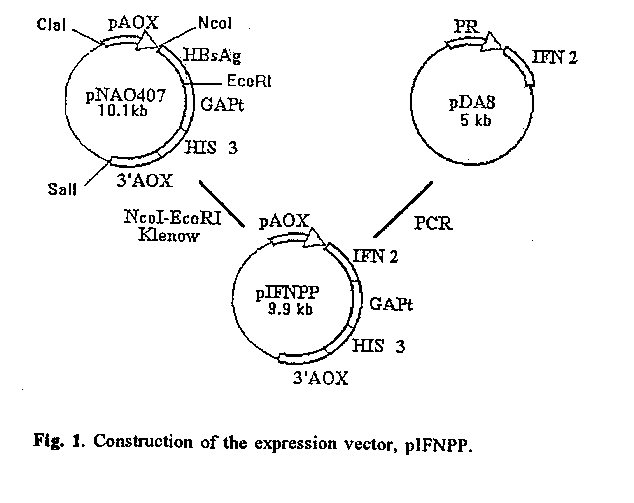
RESULTS AND DISCUSSION. Vector construction and methanol-regulated expression of IFN-alpha2 in P. pastoris The vector pIFNPP (figure 1) is expected to direct the integration of the expression cassette by homologous recombination into the P. pastoris AOX1 locus.
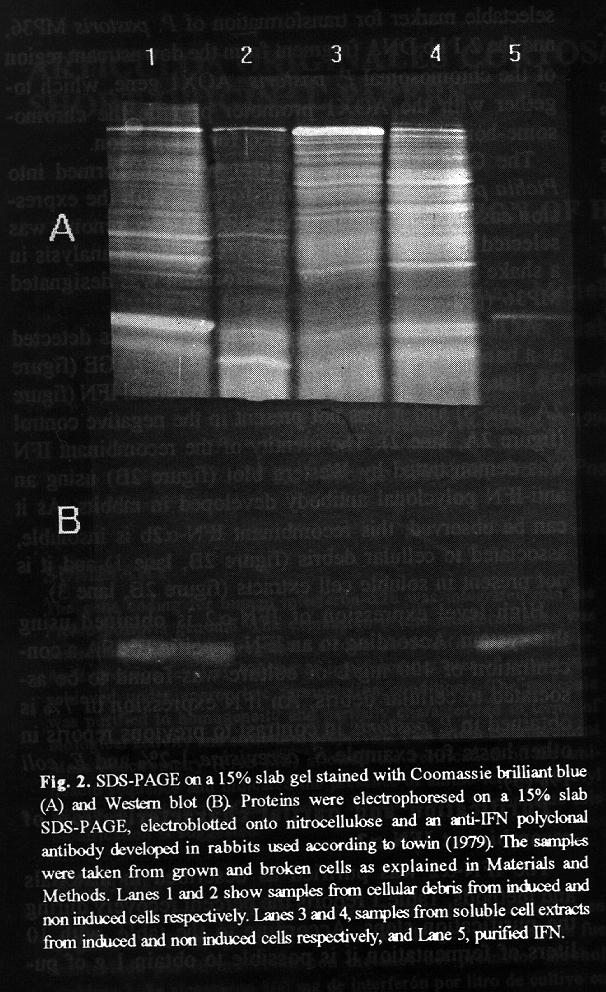
This cassette contains: the methanol regulated P. pastoris alcohol oxidase I promoter (AOX1 p), the IFN gene, S. cerevisiae GAP transcriptional termination signal (GAP t), the S. cerevisiae HIS3 gene which provided a selectable marker for transformation of P. pastoris MP36, and the 2.1 kb DNA fragment from the downstream region of the chromosomal P. pastoris AOX1 gene, which together with the AOX1 promoter provide the chromosome-homologous ends needed for integration. The ClaI-SalI digested vector was transformed into Pichia pastoris MP36. A transformant with the expression cassette integrated into the P. pastoris genome was selected by Southern blot and by expression analysis in a shake flask culture. This transformant was designated MP36 (pIFNPP). At the end of methanol induction, IFN was detected as a band in Coomassie blue-stained SDS-PAGE (figure 2A, lane 1). This band comigrated with natural IFN (figure 2A, lane 5) and it was not present in the negative control (figure 2A, lane 2). The identity of the recombinant IFN was demonstrated by Western blot (figure 2B) using an anti-IFN polyclonal antibody developed in rabbits. As it can be observed, this recombinant IFN-alpha2b is insoluble, associated to cellular debris (figure 2B, lane 1) and it is not present in soluble cell extracts (figure 2B, lane 3).
Purification and partial characterization of recombinant IFN-alpha2 The IFN-alpha2 was purified as described in materials and methods. Table 1 reports the percent recovery during the purification steps. According to the table, with 10 liters of fermentation it is possible to obtain 1 g of purified IFN-alpha2b to homogeneity. Figure 3 shows the chromatography profile of the immunoaffinity column used in the last purification step. The interferon eluted as a single peak from the immunoaffinity colum (figure 3A), in 15% SDS-PAGE the fraction number six (concentrated by ultrafiltration) gave a single band (figure 3B, lane 4) at an apparent molecular weight of 18 kD. Recombinant IFN appeared as a homogeneous species with the same molecular weight of the natural IFN-alpha2b.
Table 1 IFN-alpha2b recovery during purification steps.
----------------------------------------------------------- Step IFN-alpha2b (mg) Recovery % ----------------------------------------------------------- Fermentation 400 - Disruption 280 70 Extraction/Renaturation 140 50 Immunoaffinity 112 80 Concentrate 100 89 -----------------------------------------------------------
IFN-alpha2b was biologically active as shown by the cytopathic effect inhibition assay. The specific activity was found to be 1 x 10^7 I.U./mg of IFN, different to natural human interferon (2 x 10^8 IU/mg) (2). This difference could be explained by the high level of acetylation of the recombinant IFN. The amino terminal acetilation occurs following the incorporation of the first amino acids into the polypeptide chain, this phenomenon could be avoided by expressing the protein fused to another protein, for example ubiquitin (18). This hybrid protein could be processed in vivo by specific hydrolases in the cytoplasm of yeast (18) and the resultant product would have a non blocked N-terminus. REFERENCES 1. LENGYEL, P. (1982). Biochemestry of interferon and their actions. Annu.Rev.Biochem. 51:251-282. 2. ADOLF, G. R.; I. KALSNER; N. H. AHORN; I. MAURER-FOGY and K. CANTELL (1991). Natural human interferon-alpha2 is O-glycosylated. Biochemical Journal 276:511-518. 3. SILVA, A.; A. DE LA FUENTE; G. PEREZ; Y. QUINONES; V. JIMENEZ; L. NOVOA and L. HERRERA (1988). Altos niveles de expresiOn del IFN-alpha2 humano bajo el control del promotor derecho del fago lambda en E.coli. Interferon y Biotecnologia 5: 40-46. 4. CREGG J. M.; T. S. VEDVICK and W. C. RASCHKE (1993). Recent advances in the expression of foreing genes in Pichia pastoris. Biotechnology 11:905-910. 5. YONG, V.; M. E. GONZALEZ; L. HERRERA and J. DELGADO (1992). HIS-3 gene of Saccharomyces cerevisiae complement his mutation in yeast Pichia pastoris. BiotecnologIa Aplicada 9:55-61. 6. SAMBROOK, J.; E. F. FRITSCH and T. MANIATIS (1989). Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, New York. 7. GALZY, P. and P. P. SLONIMSKY (1957). Variation phisiologique de la levure au coure de la croissauce sur 1`acide lactique comme seule source de carbone. Comptes. Rend. Acad. Sci. (Paris) 245:2423-2433. 8. JIMENEZ, V.; G. PADRON; L. CASTELLANOS y L. RODES (1984). Sintesis quimica de oligonucleotidos por el metodo de fosfodiester. Interferon y Biotecnologia 1:39-57. 9. SAIKI, R.; S. SCHARF; F. FALOONA; K. B. MILLES; G. T. HORN; H. A. ERLICH and N. ARNHEIM (1985). Enzymatic amplification of beta-globin genomic sequences and restriction site analysis for diagnosis of sickle cell anemia. Science 230: 1350-1354. 10. HINNEN, A.; J. B. HICK and G. R. FINK (1978). Transformation of yeast. Proceedings of the Nattional Academy of Sciences USA 75:1929-1933. 11. CRUZ, S.; C. DUARTE; E. FERRA; G. FONTIRROCHE; >J. VAZQUEZ; L. MARTINEZ; N. ARTEAGA; E. PEREZ and J. GAVILONDO (1990). Quantitation of recombinant human alpha 2b interferon with monoclonal antibodies. Biotecnologia Aplicada 7:132-141. 12. BRADFORD, M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochem. 72: 248-254. 13. Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680-685. 14. TOWIN, H.; T. STAEHLIN and J. GORDON (1979). Electrophoretic transfer of proteins from polyacrilamide gels to nitrocellulose sheets. Proceedings of the National Academy of Sciences USA 76: 4350-4354. 15. PEREZ, E.; V. MARANON; L. PEREZ (1992). Tratamientos analiticos para muestras de IFN-alpha2b humano recombinante para su cuantificacion por ELISA. Biotecnologia Aplicada 9:1. 16. BARCELONA, S. (1983). Determinacion de la actividad de interferon. En: Memorias I Seminario Cubano sobre Interferon, Ed. Inst. de Cooperacion Iberoamericana, 47-51. 17. BESADA, V.; W. ANTUCH; R. RODRIGUEZ; L. GONZALEZ; G. CHINEA; G. PADRON (1992). Mass spectrometry in protein analysis. Biotecnologia Aplicada 9:1-15. 18. JENTSCH, S.; W. SEUFERT and H. P. HAUSER (1991). Genetics analysis of the ubiquitin system. Biochemica et Biophysica Acta 1089: 127-139. Copyright 1995 Sociedad Iberolatinamericana de Biotecnologia Aplicada a la Salud
The following images related to this document are available:Photo images[ba95052b.jpg] [ba95052c.jpg]Line drawing images[ba95052a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}