 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
ARTICULO SOBRE TECNICA / PAPER ON TECHNIQUE CLONING AND EXPRESSION OF GENES CODING FOR Haemophilus influenzae Rf RESTRICTION AND MODIFICATION ENZYMES. PURIFICATION OF THE RECOMBINANT ENDONUCLEASE Arlene Rodriguez^1, Ricardo Lleonart^2, Ana S. Martinez^3, Geovani Reyes^4, Ernesto Gonzalez^5 and Jose E. Brito^3 ^1Division de Farmaceuticos. Centro de Investigaciones Biologicas. P.O. Box 6996. Ciudad de La Habana, Cuba. ^2Division Genetica de Celulas de Mamiferos. Centro de Ingenieria Genetica y Biotecnologia. P.O. Box 6162. La Habana 6, Cuba. ^3 Division de Desarrollo Biofarmaceutico.C.I.G.B, P.O. Box 6162. La Habana 6, Cuba. ^4Division de Produccion de Reactivos Biologicos. C.I.G.B, P.O. Box 6162. La Habana 6, Cuba. ^5Division de Quimica-Fisica. C.I.G.B, P.O. Box 6162. La Habana 6, Cuba. Recibido en octubre de 1994. Aprobado en mayo de 1995
Key Words: Hinf I, Methylase, Restrictase, Restriction-Modification System.
SUMMARY The genes coding for Hinf I restriction and modification enzymes were cloned from the Haemophilus influenzae Rf strain using pUC18 plasmid as vector. To select the genes from the library, the classical modification phenotype selection was used. When introduced into Escherichia coli HB101 strain, the recombinant plasmid pERHinf4 was able to direct the expression of the restriction phenotype. The crude extracts from the transformed E. coli contained roughly twice the amount of restrictase expressed by the natural source. A purification procedure for the restrictase is described which when applied to the recombinant enzyme, renders a preparation free of contaminant exonucleases and phosphatases. The overall recovery of the purification procedure was about 33% of the total activity.
RESUMEN Utilizando el clasico metodo de seleccion para el fenotipo de modificacion, se aislaron los genes que codifican para la restrictasa y la metilasa Hinf I. Se utilizo el plasmido pUC18 para la construccion de la libreria genomica, asi como para la expresion de estas proteinas en Escherichia coli. El clon obtenido a partir del proceso de seleccion (pERHinf4) mostro expresion de la restrictasa en la cepa de E. coli HB101. Los extractos celulares obtenidos a partir de esta cepa transformada mostraron el doble de la cantidad (U/g de biomasa) de restrictasa que usualmente contienen los de la cepa Haemophilus influenzae Rf. En este trabajo describimos un procedimiento de purificacion de esta restrictasa recombinante expresada en E. coli. Este procedimiento permite obtener una preparacion libre de fosfatasas y exonucleasas contaminantes, con un recobrado final de enzima equivalente al 33% de la actividad total presente en el extracto.
INTRODUCTION Several restriction and modification enzymes have been cloned from Haemophilus sp. (Wilson, 1988b; Slatko et al., 1988). The purification of the Hinf I endonuclease was undertaken from a recombinant source because of the high cost of purification from natural sources. The enzyme yield of the purification procedure from the natural source is approximately 160 U/g of wet cells and the quality of the enzymatic preparation obtained is very variable because of the presence of exonuclease and phosphatase activities. The molecular cloning of the Hinf I restriction and modification enzymes coding genes has already been reported (Chandrasegaran et al., 1988). These authors increased the expression levels in extracts from 150 U/g of wet cells to 300 000 by using a multicopy vector. The cloning of the Hinf I RM (Restriction and Modification) genes based on selection for the modification phenotype (Lunnen et al., 1988; Wilson, 1988a; Chandrasegaran et al., 1988) is reported here. The restrictase was expressed in E. coli HB101 strain at a wvel of 50 000 U/g of wet cells. A purification procedure is described for the recombinant enzyme which allows an overall recovery of 33%. The enzyme preparations obtained from this protocol were free of spurious phosphatase and exonuclease activities.
MATERIALS AND METHODS Bacterial strains, plasmids and culture conditions The Haemophilus influenzae Rf strain was obtained from the C.I.G.B. collection (No.49). It was grown in an orbital shaker, using a growth medium containing, per liter, 10 g of casein hydrolyzate, 5 g of yeast extract, 4.4 g of K2HPO4, 2 g of NaCl, 2 g of dextrose, 2 mg of NAD and 10 mg of haemin. The E. coli strain HB101 [F^-, delta(gpt-proA)62 leu supE44, ara14 gal K2 lacY1 delta(mcrC-mrr) rpsL20 (Str^r) xyl-5 mtl-1 recA13] (Maniatis et al., 1982) transformed with the plasmid pERHinf4 (described at Results and Discussion section) was grown in an LBA medium containing, per liter, 10 g of tryptone, 5 g of yeast extract, 10 g of NaCl and 50 ug/mL of ampicillin. Cells were harvested when OD600 = 0.8 by centrifugation at 4 000 x g for 30 min at 4 C, and stored at -20 C. Enzymes and reagents Restriction and modification enzymes were obtained from HeberBiotec SA (Havana, Cuba). Calf intestinal phosphatase (CIP) was obtained from Boehringer Manheim (Germany). Enzymes were used according to the manufacturer's recommendations. All reagents used were of analytical grade quality. Construction of a Haemophilus influenzae Rf genomic library in pUC18 High molecular weight DNA was isolated from Haemophilus influenzae Rf (Caserta et al., 1987) and cleaved completely with Pst I endonuclease. The region between 2.3 and 4.3 kb was purified from agarose gels and ligated into a pUC18 vector prepared by digestion with Pst I and alkaline phosphatase treatment. After transformation into E. coli HB101, each plate was flooded with an LBA medium and the colonies were scraped together and pooled. Cells were harvested and plasmids were extracted by the alkaline-SDS procedure and further purified by precipitation with polyethylene glycol (Maniatis et al., 1982). Selection for the modification phenotype Plasmid DNA from the library (4 ug) was digested to completion at 37^oC for 1 hour with 10 units of Hinf I endonuclease and E. coli HB101 was transformed with the product of the digestion. A new pool of plates was done and their plasmids were purified and digested again with Hinf I endonuclease as explained before. Surviving colonies were individually picked up and cultured overnight in 5 mL of LBA medium. Cells were harvested and plasmids were purified by the miniprep procedure (Maniatis et al., 1982). Plasmids were analyzed for resistance to Hinf I cleavage and for the presence of insert. Hinf I restriction endonuclease assay Escherichia coli cultures harboring the recombinant plasmids were grown overnight in 300 mL of LBA medium. Cellular extracts were prepared by sonication, and incubated with 1 ug of lambda DNA in the presence of 50 mM NaCl, 10 mM TrisHCl pH 7.9, 10 mM MgCl2, 1 mM dithiothreitol for 1 hour at 37^oC in a final volume of 50 uL. One unit of activity was defined as the amount of enzyme required to cleave the substrate to completion under the conditions described. The products of the digestion with different amounts of extract were analyzed by ethidium bromide staining of agarose gels. Protein determinations Protein concentrations were determined as described by Bradford (Bradford, 1976). The method is based on the interaction between the Commasie blue reagent and the peptidic bridge of the proteins. Purification of recombinant Hinf I endonuclease from E. coli HB101 transformed cells A kinetic study was performed to determine the optimal O.D. for harvesting the E. coli cultures containing the pERHinf4, concerning the production of the restrictase. After harvesting the cells by centrifugation at 4 C, the following procedure was performed. The cell paste (6 g wet weight) was suspended in 12 mL of the PC buffer (10 mM KPO4, 10 mM 2-mercaptoethanol, 0.1 mM EDTA, pH 7) with 0.5 mM PMSF, 1 mM benzamidine and disrupted in the French press. The lysate was clarified by centrifugation at 27 000 x g for 2 h at 4 C. The supernatant was precipitated using ammonium sulfate (70%) and centrifuged at 12 000 x g for 10 min at 4 C. The pellet was suspended and dialyzed for 12 h against the PC buffer with 150 mM NaCl. The extract was loaded onto a Phosphocellulose P-11 (Whatman, 2.6 x 40 cm column) with a bed volume of 100 mL, previously equilibrated with the PC buffer containing 150 mM NaCl, at a flow rate of 25 mL/h. The column was washed with 1 L of the PC buffer with 150 mM NaCl and eluted with a linear gradient of NaCl (0.15-0.8 M). The Hinf I activity was eluted between 0.3 and 0.49 M NaCl. Pooled fractions were dialyzed against the PC buffer for 4 h and then loaded onto a Heparin-Sepharose column (Pharmacia, 1 x 10 cm) with a bed volume of 10 mL, previously equilibrated with the PC buffer, at a flow rate of 20 mL/h. The column was washed with 100 mL of the PC buffer and eluted with a linear gradient of NaCl (0-0.8 M). The Hinf I activity was eluted between 0.25 and 0.35 M NaCl. Fractions were pooled, concentrated with polyethylene glycol (Mw. 20 000) and dialyzed against the concentration buffer (50 mM KCl, 10 mM TrisHCl, 0.1 mM EDTA, 1 mM DTT, 50% glycerol, pH 7.4). This preparation was stored at -20 C. Phosphatase and exonuclease assays The presence of phosphatase and exonuclease contaminating activity in the final preparation of recombinant and natural Hinf I endonuclease was performed, as previously described (Gonzalez, 1990).
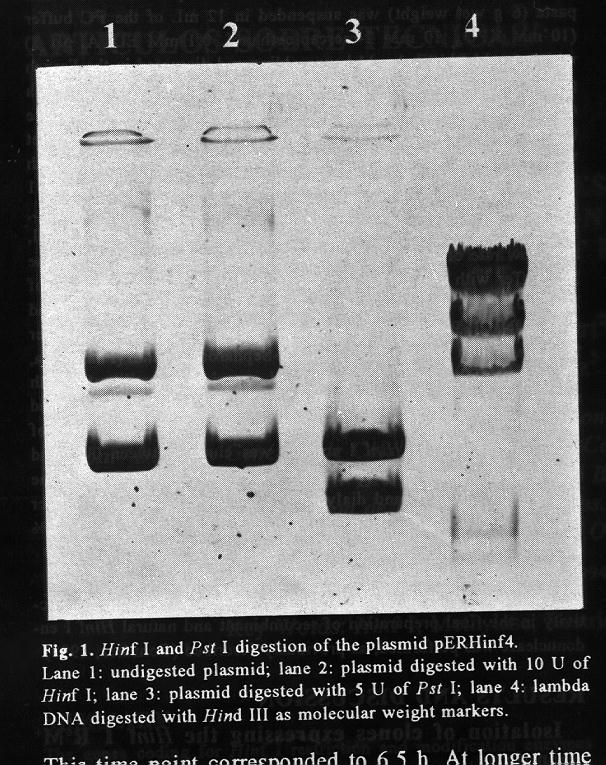
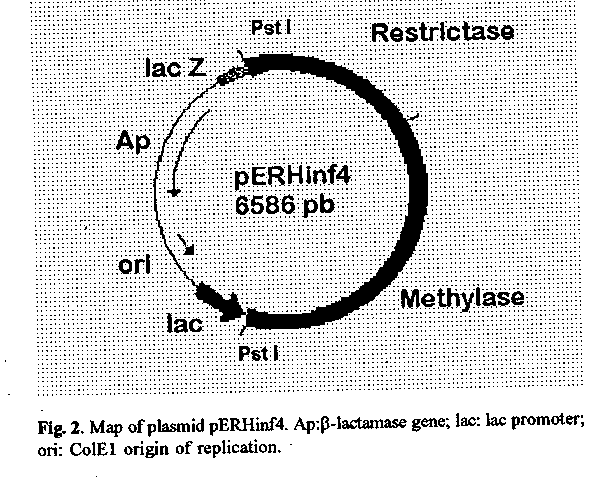
RESULTS AND DISCUSSION Isolation of clones expressing the Hinf I R^+M^+ phenotype Approximately 10 000 recombinant colonies from the Pst I library were screened for the modification phenotype. When plasmid DNA from the library was overdigested with Hinf Y and transformed into E. coli, a population of resistant colonies was obtained. After a second round of overdigestion and transformation, a few hundred resistant colonies were obtained. One of the recombinant plasmids, the pERHinf4, contained the 3.9 kb insert (figure 1 and 2) corresponding to the chromosomal fragment which harbors the methylase and restrictase genes (Chandrasegaran et al., 1988).
Fig. 1 Hinf I and Pst I digestion of the plasmid pERHinf4. Lane1: undigested plasmid; lane 2: plasmid digested with 10 U of Hinf I; lane 3: plasmid digested with 5 U of Pst I; lane 4: lambda DNA digested with Hind III as molecular weight markers.
Fig.2 Map of plasmid pERHinf4. Ap: beta-lactamase gene; lac: lac promoter; ori: ColE1 origin of replication.
This plasmid was selected to assay the expression of Hinf I endonuclease activity. The extract prepared from 300 mL overnight cultures was found to contain detectable Hinf I endonuclease activity (figure 3).
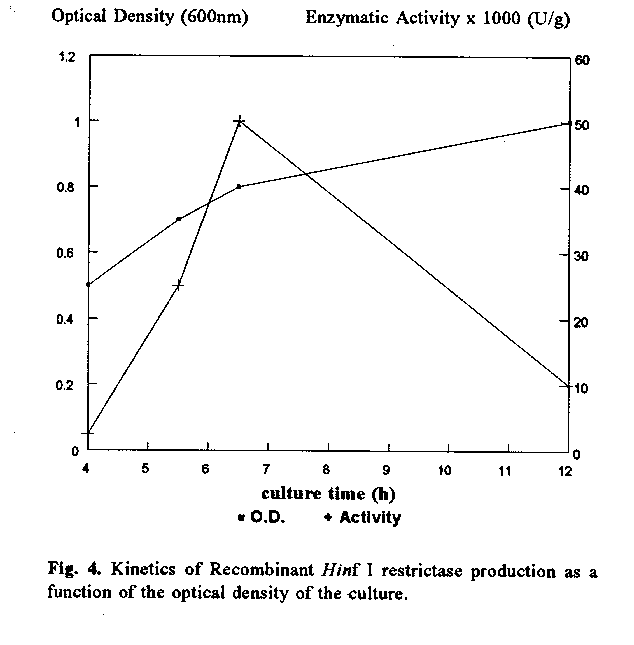
The extract prepared from 300 mL of transformed E. coli cultures expressed at least 10 000 U/g of wet cells (figure 3). When the culture was stopped at the optimal O.D. calculated from the kinetic study, a yield of about 50 000 U/g of wet cells was obtained (figure 4). This time point corresponded to 6.5 h. At longer time points, the enzymatic activity was masked by the presence of non-specific nuclease activities.
Purification of recombinant Hinf I restrictase from E. coli To the author's knowledge, there is no reported procedure for the purification of the Hinf I restrictase. Here is shown a purification procedure which allows the obtainment of an enzymatic preparation free of exonuclease and phosphatases activities. Table 1 shows the results of the different purification steps when the recombinant restrictase was purified from E. coli extracts. The final specific activity achieved was always around 49 000 U/mg. The overall yield was about 19 000 U/g of wet cells, as opposed to the very low yield (160 U/g) obtained when the restrictase was purified from H. influenzae extracts using the procedure described in Materials and Methods for the recombinant enzyme.
Table 1 Summary of the recombinant Hinf I restrictase purification procedure.
-----------------------------------------------------------
Fraction Total Total Specific Yield Purif.
activity protein activity activity (fold)
[U] [mg] [U/mg]* [%]** ***
---------------------------------------------------------
Crude
extract 300000 424 706+/-177 100 1
P-11 112000 3.3 33939+/-2785 37+/-6 48+/-15
Heparin
Sepharose 110000 2.2 49107+/-15783 36+/-12 69+/-9
-----------------------------------------------------------
Values shown are the average of three different purification processes +/- SD. * Referred as the ratio total enzymatic activity/total protein ** Referred as percentage of remaining total enzimatic activity compared with the initial amount in the crude extract. *** Purification fold calculated as the increase in specific activity compared with the crude extract. The purification of the recombinant enzyme allowed a much cleaner final preparation, without the large amounts of phosphatases and exonucleases usually present at the end of the procedure when natural extracts where employed. The use of this modified E. coli as a Hinf I source is very convenient taking into consideration, 1) the very tricky culture conditions required by the H. influenzae, 2) the higher levels of expression, and 3) the quality of the final preparation, which was devoid of undesirable activities. This preparation is suitable for molecular biology applications, as demonstrated by its continuous use in our laboratory.
ACKNOWLEDGMENTS The authors would like to express their thanks to G. Ferbeire for helpful suggestions during the course of this work and J. de la Fuente for critically reading this manuscript.
REFERENCES 1. BRADFORD, M. M. (1976). A rapid and sensitive method for the quantitation of protein utilizing the principle of protein-dye binding. Biochemistry 72: 248-254. 2. CASERTA, M.; W. ZACHARIAS; D. NWANKWO; G. G. WILSON and R. D. WELLS (1987). Cloning, sequencing, in vivo promoter mapping, and expression in E. coli of the gene for the Hha I methyltranferase. J. Biol. Chem. 262: 4770-4777. 3. CHANDRASEGARAN, S.; K. D. LUNNEN; H. O. SMITH and G. G. WILSON (1988). Cloning and sequencing the Hinf I restriction and modification genes. Gene 70: 387-392. 4. GONZALEZ, E.; J. BRITO; L. TRUJILLO; M. ORTEGA; M. VAZQUEZ; Y. JORGE and O. GARCIA (1990). Quality controls of enzymes restriction and/or modification enzymes. In: Book of short reports of the 2^nd Latioamerican Congress of Biotechnology. August , Havana, Cuba. 5. LUNNEN, K. D.; J. M. BARSOMIAN; R. R. CAMP; C. O. CARD; S. Z. CHEN; R. CROFT; M. C. LOONEY; M. M. MEDA; L. S. MORAN; D. O. NWANKWO; B. E.SLATKO; E. M. VAN COTT and G. G. WILSON (1988). Cloning type II restriction and modification genes. Gene 74: 25-32. 6. MANIATIS, T.; E. FRISCH and J. SAMBROOK (ed.) (1982). Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York. 7. SLATKO, B. E.; R. CROFT; L. S. MORAN and G. G. WILSON (1988). Cloning and analysis of the Hae III and Hae II methyltranferase genes. Gene 74:45-50. 8. WILSON, G. G. (1988a). Type II restriction-modification systems. Trend in Genetics 4:314-318. 9. WILSON, G. G. (1988b). Cloned restriction-modification systems - a review. Gene 74:281-289.
Copyright 1995 Sociedad Iberolatinamericana de Biotecnologia Aplicada a la Salud
The following images related to this document are available:Photo images[ba95053a.jpg] [ba95053c.jpg]Line drawing images[ba95053d.gif] [ba95053b.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}