 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Carbohydrate features of recombinant human glycoproteins Johannis P Kamerling
Bijvoet Center, Department of Bio-Organic Chemistry, Utrecht
University, P.O. Box 80.075, NL-3508 TB Utrecht, The
Netherlands.
Code Number:BA96073
Size of Files:
Text: 68.7K
Graphics: Line drawings (gif) - 93.1K
Abstract In the present approaches of developing pharmaceutically relevant drugs based on recombinant human glycoproteins, the establishment of the glycosylation patterns of these biomolecules is highly actual. In this minireview a short overview of the state of the art with respect to glycoprotein glycan analysis is given. Furthermore, the glycosylation of plasminogen activators and erythropoietins is summarized in more detail. Key words: recombinant glycoproteins, carbohydrate analysis, plasminogen activator, erythropoietin Resumen El establecimiento de los patrones de glicosidacion de farmacos basados en glicoproteinas humanas recombinantes constituye un elemento de gran actualidad en las tendencias modernas de desarrollo de esas biomoleculas. El presente trabajo de revision pasa revista al estado del arte del analisis de glicanos de las glicoproteinas. Ademas, sumariza en mayor detalle la glicosidacion de los activadores del plasminogeno y las eritropoietinas. Palabras claves: glicoproteinas recombinantes, analisis de carbohidratos, activador de plasminogeno, eritropoietina Introduction In recent years the academic and industrial interest in the carbohydrate part of glycoproteins has grown dramatically. This is mainly due to accumulating evidence concerning the relevance of the glycan chains in the physical and biological behaviour of these biomolecules. N-linked glycoprotein glycans award essential physical properties like proper folding and stabilization of the conformation of glycoproteins, protease resistance, and charge and waterbinding capacity. Furthermore, they are relevant in biological recognition processes like protein targetting and cell-cell interactions. It has been well established that ensembles of N-linked carbohydrate chains of glycoproteins, especially the range of non-reducing terminal structural elements, are species specific, tissue specific, cell-type specific, and protein specific. The terminal sequences are often differentially expressed during early development, provide masking functions to prevent rapid clearance from circulation, activate host immunological systems, act as immunodeterminants, and influence bioactivity. The importance of O-linked carbohydrate chains has been mainly discussed in terms of charge, waterbinding properties, and stability. For comprehensive reviews, see Rademacher, et al. (1), Allen and Kisailus (2), Lis and Sharon (3) and Mon-treuil, et al. (4,5).
It is without doubt that the present knowledge on the importance of glycoprotein glycosylation has strongly influenced the discussion with respect to genetically engineered proteins prepared in heterologous cell types, with or without a glycosylation machinery (6-8). In the context of therapeutic administration of recombinant human glycoproteins, an increasing interest of pharmaceutical industries is observed to consider the glycosylation patterns of the engineered proteins, with regard to applicability, tolerance and patent position.
The development of advanced analytical methods to unravel the structures of the ensembles of carbohydrate chains, present at specific amino acid residues in polypeptide backbones, has been the first step in the glycoprotein revival, and formed the basis for what today is called "glycobiology". Although in recent years several approaches based on ^1HNMR spectroscopy (9, 10) and mass spectrometry (11-16) have become available, and much research is focused on the development of profiling techniques based on high performance separation procedures (17-19), the primary structure analysis of glycoprotein glycans has still not reached the level of a routine analysis. Even now, completely novel oligosaccharide elements, as part of glycoprotein glycans, are identified. Therefore, it should be stressed that, due to the complexity of the carbohydrate chains, not only in terms of monosaccharide constituents and sequences, but also in terms of non-carbohydrate substituents, the current approaches have to be applied with great care.
In this minireview, the chemical nature of glycoprotein glycans and the current methodologies for the analysis of glycosylation patterns will be shortly described. Special attention will be paid to the glycosylation of two pharmaceutically important glycoproteins, namely, human plasminogen activator and human erythropoietin. Structural Aspects of Glycoprotein Glycans Glycoproteins are biopolymers consisting of a polypeptide backbone with covalently attached carbohydrate chains (2-5). Two major classes occur, namely, N-glycans linked via N-acetyl-beta-D-glucosamine (GlcNAc) to the amide nitrogen of L-asparagine (Asn), and O-glycans linked via N- acetyl-alpha-D-galactosamine (GalNAc) to the hydroxyl group of L-serine (Ser) or L-threonine (Thr). Glycoproteins may contain more than one glycosylation site, and N- as well as O-linked glycans can be attached to one polypeptide backbone. Moreover, microheterogeneity of the oligosaccharide structures attached to one amino acid residue is a known phenomenon, yielding a series of distinct glycoforms for each glycoprotein.
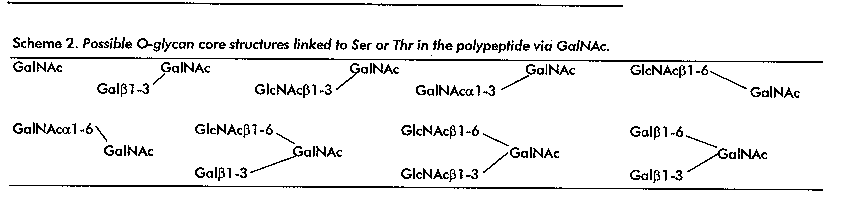
For the mentioned O-glycans, so far nine core elements can be defined, which are depicted in Scheme 2. These cores can be extended in several ways, comprising e.g. Galbeta1-4GlcNAcbeta or Galbeta1-3GlcNAcbeta (Gal, D-galactose) elements, and terminated by monosaccharides like L-fucose (Fuc), Gal, GalNAc or N-acetylneuraminic acid (Neu5Ac) / N-glycolylneuraminic acid (Neu5Gc) (10). Moreover, O-acetyl and sulfate groups can occur, as demonstrated for the N-glycans.
Structure Analysis of Glycoprotein Glycans
----------------------------------------------------------
Scheme 3. Major methodologies in the structure
analysis of glycoprotein glycans. PNGase,
peptide-N4-(N-acetyl-beta- glucosaminyl)-asparagine amidase;
PA, 2-pyridylamino; LC, liquid chromatography; GLC-MS,
gas-liquid chromatography / mass spectrometry; 1D, one
dimensional; 2D, two dimensional; NMR, nuclear magnetic
resonance.
-------------------------------------------------------------
* Isolation of glycoproteins
* Preparation of carbohydrate chains
- protease digestion ---> glycopeptides
- hydrazinolysis far GlcNAc-Asn type ---> oligosaccharides /
alditols / PA derivatives
- PNGase F or A far GlcNAc-Asn type ---> oligosaccharides /
alditols / PA derivatives
- endo-H and other endo-N-acetylglucosaminidases
far GlcNAcAsn type (mainly ---> oligosaccharides /
oligomannose type) alditols /
PA derivatives
- alkaline borohydride treatment ---> oligosaccharide-
for GaINAc-Ser/Thr type alditols
* Fractionation and purification,
if possible to homogeneity
- gel-permeation chromatography ---> high-voltage paper
(Bio-Gel P-4) electrophoresis
- high-pH anion-exchange --->lectin chromatography
chromatography ICarboPac)
- high-pressure chromatography ---> fast-protein liquid
(e.g. Lichrosorb-NH2) chromatography (Mono Q)
* Degradation procedures
- exo-glycosidases (alpha-sialidases, alpha-D-
galactosidases, beta-D-galactosidases,
N-acetyl-beta-D- hexosaminidases,
alpha-D-glucosidase, alpha-D-
mannosidases, beta-D-mannosidases, alpha-L-fucosidases,
beta-D-xylosidase)
- endo-glycosidases (endo-beta-D-galactosidases)
- partial solvolysis
- periodate oxidation
* Monosaccharide analysis, including absolute configuration
determination (GLC-MS)
* Methylation analysis (GLC-MS)
* 1 D/2D NMR spectroscopy (^1H, ^13C, ^31P)
* Mass spectrometry (electron impact, chemical ionization,
fast-atom bombardment, electrospray/ionspray, matrix-
assisted laser desorption ionization)
* Profiling (LC mapping techniques, high-pH anion-exchange
chromatography, capillary electrophoresis, fluorophore-
assisted-carbohydrate electrophoresis, reagent-array
analyses; combination with exoglycosidases or mass
spectrometry)
--------------------------------------------------------------
In view of the presented complexity of the analytical problem, it may be evident that the determination of the primary structure of Asn-linked and of Ser- or Thr-linked carbohydrate chains on intact glycoproteins is nearly impossible. Therefore, the preparation of partial structures of the protein, having a single glycosylation site (glycopeptides), or release of the glycan chains from the glycoprotein (oligosaccharides), is a prerequisite. In Scheme 3 the major cleavage procedures for the preparation of mixtures of glycopeptides or oligosaccharides from native glycoproteins are summarized (2, 4, 5, 17-21). For a detailed analysis of the released carbohydrate chains, these mixtures have to be fractionated, and different combinations are in use. Frequently applied combinations are high-voltage paper electrophoresis and Bio-Gel P-4 gel-permeation chromatography (19, 20, 22), or Mono Q/Resource Q, Lichrosorb-NH2 and CarboPac liquid chromatography (23, 24). For additional information, see also Fukuda and Kobata (17), Hounsell (18) and Lennarz and Hart (19). The structural analysis of isolated carbohydrate chains can be carried out along different routes. Major strategies include methylation analysis (12), exo- and endo-glycosidase studies (2, 17-20), ^1H-NMR spectroscopy (9, 10) and mass spectrometry (11, 13, 15, 16). Quite often these approaches are used in combination with each other. But also other approaches have been reported (for general reviews, see Fukuda and Kobata (17), Hounsell (18) and Lennarz and Hart (19). At present, great effort is placed on the development of profiling procedures, which can be applied to get an impression of the carbohydrate chains present in unknown glycoproteins, or will be of use in controlling possible glycan microheterogeneity shifts in known glycoproteins (batch control). In this respect, more-dimensional HPLC mapping techniques (25), high-pH anion-exchange chromatography (26, 27), capillary electrophoresis (28, 29), fluorophore-assisted- carbohydrate electrophoresis (30), and so-called reagent-array analyses (31) seem to be highly promising.
Over the years, part of our research projects have been focused on the glycan analysis of a wide range of glycoproteins derived from many different biological sources. For an idea of our activities in this field during the last five years, see Bergwerff, et al. (32-35), Damm, et al. (36), De Waard, et al. (37), Gerwig, et al. (38), Hard, et al. (23, 39-41), Hokke, et al. (24, 42, 43), Kubelka, et al. (44), Pfeiffer, et al. (45), Van Dam, et al. (46) and Van Zuylen, et al. (47). Recombinant Human Glycoproteins Several reports have appeared in the literature focusing on primary structure analysis of recombinant glycoproteins and on the biological significance of the specific glycosylation patterns. Typical examples of the structure analysis of recombinant human glycoprotein glycans, as carried out in our research group, comprise the carbohydrate chains of gamma-interferon (N-glycoprotein; 48), follitropin (N-glycoprotein; 39), chimeric plasminogen activator (N-glycoprotein; 32) and erythropoietin (N,O-glycoprotein; 43, 49, 50), all produced in Chinese hamster ovary (CHO) cells. Furthermore, detailed studies were carried out on the glycan chains of IgG1 antibodies (N-glycoprotein; 34), produced in murine SP2/0 transfectoma cell subclones, and of insuline-like growth factor I (O-glycoprotein; 51), produced in Saccharomyces cerevisiae cells. In all these cases, ^1H-NMR spectroscopy was applied as the major analytical technique. Plasminogen Activators Plasminogen activators (PA) are serine proteases, that catalyze the conversion of inactive plasminogen into active plasmin (52-54). Plasmin is considered to be the primary circulating fibrinolytic enzyme, which can clear thrombi from blood vessels by the degradation of the fibrin network, a process called fibrinolysis. In view of this feature, PA's are of clinical interest for the treatment of thrombolytic disorders. Two major activators have been identified, namely, tissue-type plasminogen activator (t PA) and urinary-type plasminogen activator (u PA). Besides the generation of recombinant plasminogen activators, hybrid variants have also been constructed, and in the framework of this minireview one of these chimeric plasminogen activators, K2tu-PA, is included. Tissue-Type Human Plasminogen Activators The polypeptide backbone of human t-PA, having a Mr of about 70 kDa and built up from 527 amino acids (an N-terminal extension of 3 amino acids not included), consists of an N-terminal finger (F) domain, an epidermal growth factor (E) domain, two kringle (K1 and K2) domains, and a C-terminal pro- tease (P) domain (54). It exists in a single-chain form and in a two-chain form, and the single-chain form can be converted into the two-chain form by the action of proteases like plasmin (cleavage between Arg-275 and Ile-276). In the two- chain form an A-chain comprises the domains F, E, K1 and K2, and a B-chain correlates with the P domain; the two polypeptides are linked through a disulfide bridge. Several reports have discussed the influence of the glycosylation of t-PA on its biochemical / biological properties, and specific information can be obtained from Hansen, et al. (55), Beebe and Aronson (56), Collen, et al. (57), Lucore, et al. (58), Wittwer, et al. (59), Wittwer and Howard (60), Howard, et al. (61), Otter, et al. (62), Berg, et al. (63) and Hajjar and Reynolds (64).
--------------------------------------------------------------
Table 1. Survey of structural information concerning major
N-glycosylation patterns of human tissue-type plasminogen
activators.
--------------------------------------------------------------
N-glycosylation sites
------------------------------------------
Cell Type Asn-117
--------------------------------------------------------------
Human colon predominantly oligomannose type;
fibroblast cells (65)
Bowes melanoma cells almost exclusively oligomannose type
(67)
Chinese hamster ovary almost exclusively oligomannose type
cells (49, 67, 70)
C127 mouse epithelial predominantly oligomannose type
cells (45, 71)
N-glycosylation sites
------------------------------------------
Cell Type Asn-184 (+/-)
--------------------------------------------------------------
Human colon predominantly N-acetyllactosamine type;
fibroblast cells (65) NeuSAc in alpha-2-6 linkage
Bowes melanoma cells predominantly N,N'-diacetyllactose-
(67) diamine type; Neu5Ac in alpha-2-6
linkage
Chinese hamster ovary predominantly N-acetyllactosamine type;
cells (49, 67, 70) Neu5Ac in alpha-2-3 linkage; traces of
NeuSGc
C127 mouse epithelial N-acetyllactosamine type; Neu5Ac in
cells (45, 71) alpha-2-3/6 linkage; NeuSAc alpha-
2-3(SO4-6)-Gal; Gal-alpha1-3Gal
N-glycosylation sites
------------------------------------------
Cell Type Asn-448
--------------------------------------------------------------
Human colon predominantly N-acetyllactosamine
fibroblast cells (65) NeuSAc in alpha-2-6 linkage
Bowes melanoma cells predominantly N,N'-diacetyllactose-
(67) diamine type; Neu5Ac in alpha-2-6
linkage
Chinese hamster ovary predominantly N-acetyllactosamine
cells (49, 67, 70) type; Neu5Ac in alpha-2-3 linkage;
traces of NeuSGc
C127 mouse epithelial N-acetyllactosamine type; Neu5Ac
cells (45, 71) in alpha-2-3/6 linkage; NeuSAc
alpha-2-3(SO4-6)-Gal;Gal-alpha-1-3Gal
--------------------------------------------------------------
t-PA is N- and O-glycosylated, and has a carbohydrate content of approximately 7 %. Two variants of t-PA occur, namely, type I t-PA, which is N-gly-cosylated at Asn-117 (K1 domain), Asn-184 (K2 domain), and Asn-448 (P domain), and type II t-PA, which is glycosylated at Asn-117 and Asn-448. A Fuc residue is O-linked to Thr-61 in the E domain. So far, the primary structure of the human serum t-PA N-glycans has not been determined, but studies addressing the elucidation of the glycan chains of non-recombinant t-PA from human colon fibroblast cells (65) and Bowes melanoma cells (65-67), and of recombinant human t-PA from CHO cells (49, 68-70) and C127 mouse epithelial cells (45, 69, 71) have been reported. An overview of the results is presented in Table 1. An evaluation of the various studies shows that each cell type produces t-PA with a unique glycosylation pattern, wherein oligomannose-, N-acetyllactosamine-, N,N'-diacetyllactosedia-mine- and/or hybrid-type structures can play a role. Of special interest are the differences in the type of linkage for sialic acid (alpha2-6 versus alpha2-3), the predominant/ almost exclusive glycosylation of Asn-117 with oligomannose-type structures, the exclusive occurrence of N,N'-diacetyllactosediamine-type structures in t-PA from Bowes melanoma cells, and the exclusive sulphated structure in recombinant t-PA from C127 mouse epithelial cells (Scheme 4). In some of the studies the detection of oligo- (N-acetyllactosamine) elements was mentioned. The different studies focused on t-PA from Bowes melanoma cells clearly illustrate the complexity of glycan analysis.
Comparison of the glycan ensembles of type I and type II t-PA showed that the presence (type I) or absence (type II) of carbohydrate chains at Asn-184 did not affect the glycosylation patterns at Asn-117 or at Asn-448 (70). Furthermore, a recombinant t-PA from CHO cells bearing a single N-glycosylation site at Asn-448 (Gln in stead of Asn at positions 117 and 184) demonstrated the only presence of N-acetyllactosamine-type chains (72). The creation of new N-glycosylation sites in the E domain of recombinant t-PA from C-127 cells afforded interesting results (73). A replacement of Tyr-67 by Asn leads to an exclusively oligomannose-type glycosylation of Asn-117, and N- acetyllactosamine-type glycosylations of Asn-67, Asn-184 and Asn-448. However, a replacement of Gly-60 by Ser leads, besides N-acetyllactosamine-type glycosylations of Asn-58, Asn-184 and Asn-448, to a predominantly N-acetyllactosamine-type glycosylation of Asn-117. For additional studies with respect to alterations in the domain structures of t-PA and the effects on glycosylation patterns, see Wilhelm, et al. (74). Interestingly, a recent study appeared dealing with t-PA coexpressed in CHO cells with alpha2-6 sialyltransferase, affording N-glycans with alpha2-6 linked sialic acid (75). Urinary-Type Human Plasminogen Activator The polypeptide backbone of human u-PA, having a Mr of about 54 kDa and built up from 411 amino acids, consists of an N- terminal epidermal growth factor (E) domain, a kringle (K) domain, and a C-terminal protease (P) domain (54). It is mainly synthesized as a single-chain form, called pro- urokinase, but for the greater part converted into a two-chain form, called urokinase, by the action of proteases like plasmin (cleavage between Lys-158 and Ile-159). In urokinase the two polypeptides (A-chain/E-K domains and B-chain/P domain) are linked through a disulfide bridge. u-PA has gained biomedical interest, since it is also involved in physiological and pathological tissue destruction and cell migration processes, such as gland involution and tumor growth (52, 76). Only limited biochemical information focused on the glycosylation is available, and can be obtained from Zamarron, et al. (77), Sarubbi, et al. (78), Henkin, et al. (79), Li, et al. (80) and Lenich, et al. (81).
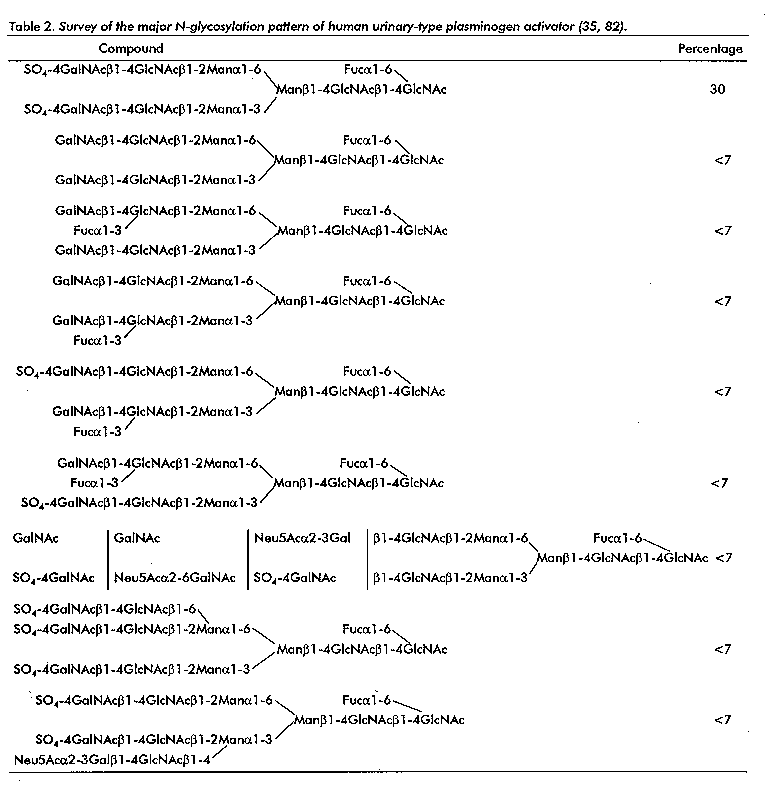
u-PA is O- and N-glycosylated, and has a carbohydrate content of approximately 2 %. A Fuc residue is O-linked to Thr-18 in the E domain. The P domain contains an N-glycosylation site at Asn-302, and recently a study describing the analysis of eleven oligosaccharides has been completed (35, 82). Detailed information has been included in table 2. The main compound (30 %) is a diantennary chain with two SO4-4GalNAcbeta1- 4GlcNAc antennae. Of further interest is the GalNAcbeta1-4(Fucalpha1-3)GlcNAc sequence, being a variant of the well-known Lewis x determinant.
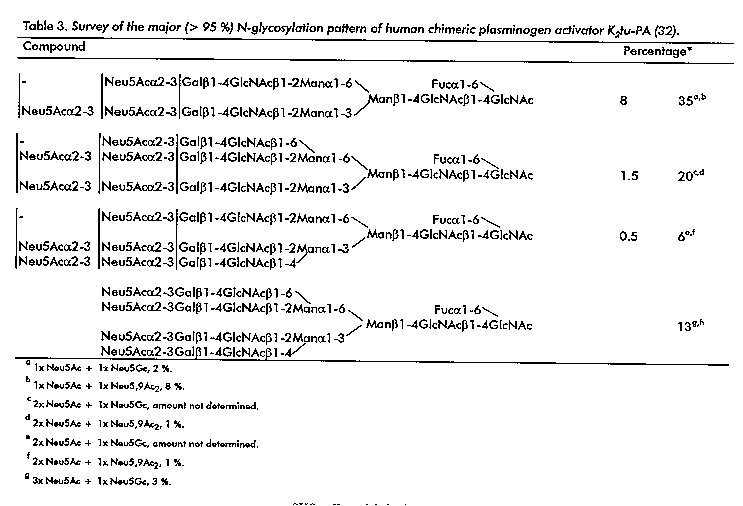
Human Chimeric Plasminogen Activator K2tu-PA In the framework of developing improved PA variants, a hybrid variant of t-PA and u-PA, namely K2tu-PA, has been constructed, wherein sequences coding for the K2 domain of t-PA and for the P domain of u-PA, were combined on the gene level, and CHO cells were chosen as expression system (83). The chimeric PA was designed in order to combine the fibrin specificity of t-PA with the catalytic activity of u-PA. K2tu-PA is partially glycosylated at Asn-12 (originally Asn-184 of t-PA) and completely glycosylated at Asn-247 (originally Asn-302 of u-PA), and has a carbohydrate content of approximately 6 %. A plasmin cleavage site, yielding the A- and B-chains, occurs between Arg-103 and Ile-104. In table 3 the structures of the various glycan chains are presented (32). They clearly represent the glycosylation machinery of CHO cells (Scheme 5). Of interest is the finding of minor amounts of Neu5Gc and Neu5,9Ac2 residues, besides the major sialic acid Neu5Ac. A discussion on the finding of Neu5Gc and the possible relevance of its appearance in recombinant human glycoproteins of pharmaceutical interest is reported by Hokke, et al. (49). The glycosylation sites on the A- (Asn-12) and the B- (Asn-247) chain show identical arrays of microheterogeneity. On the A-chain mono- and disialylated compounds have a higher relative abundance than on the B-chain, whereas the reverse holds for the tri- and tetrasialylated compounds. It should be noted that the relative amounts of the various glycan chains in K2tu-PA vary subltly with the applied isolation procedures. The highly sialylated K2tu-PA does not contain oligomannose-, hybrid-, and N,N'-diacetyllacto-sediamine-type chains. The degree of sialylation is of interest because of clearance by the hepatic Gal (asialoglycoprotein) receptor (87). The absence of oligomannose-type structures may prolong the half- life of K2tu-PA relative to that of t-PA, since the uptake from blood circulation by the hepatic Man receptor is prohibited (62). The absence of terminal SO4-4GalNAc (if present, proposed uptake by a hepatic SO4-4GalNAc receptor, (88, 89) and terminal GalNAc (if present, proposed uptake by the hepatic asialoglycoprotein receptor, (90) elements may increase the half-life of K2tu-PA relative to that of u-PA. Also the absence of O-linked Fuc in K2tu-PA may be of importance, since it has been shown that in t-PA this element may mediate binding and degradation by hepatoma cells, and it is suggested that this residue is involved in the uptake of the glycoprotein from the blood stream (64). Finally, K2tu-PA lacks the potential immunogenic determinant GalNAcbeta1- 4(Fucalpha1-3)GlcNAc.
Table 3. Survey of the major (<95 %) N- glycosylation pattern of human chimeric plasminogen activator K2tu-PA (32).
Human erythropoietin (EPO), a glycoprotein that is synthesized mainly by the kidney, stimulates the proliferation and differentiation of erythroid progenitor cells (91). The polypeptide backbone of active EPO (Mr 34 kDa) is built up from 165 amino- acids, and has three N-glycosylation sites at Asn-24, Asn-38, and Asn-83, respectively, and one O- glycosylation site at Ser-126. Its average carbohydrate content is about 40 % (92-94). Urinary EPO is generally isolated from pools of urine of aplastic anemic patients. EPO has been expressed in various heterologous cell systems and the recombinant human glycoprotein is an important therapeutic agent for the treatment of anemia associated with renal faillure (93, 95, 96). From several studies, it has become clear that the glycosylation is of great importance for the biological functioning of EPO. It could be demonstrated that removal or modification of the glycan chains, or prevention of the glycosylation at specific sites by site-directed mutagenesis, results in altered in vivo and in vitro activity (95, 97-105).
--------------------------------------------------------------
Scheme 5. Survey of structural aspeds of Chinese hamster
ovary cell derived recombinant glycoproteins.
--------------------------------------------------------------
Carbohydrate chains of recombinant human glycoproteins
expressed in CHO cells
--------------------------------------------------------------
* can contain
- (phosphorylated) oligomannose-type structures
- hybrid-type structures
- N-acetyllactosamine.type structures with mono-, di-,
tri-, tri'- and tetra-antennae, including oligo-
(N-acetyllactosamine extensions
- terminal sialic acid (Neu5Ac, Neu5,9Ac^2 and Neu5Gc),
but only in alpha-2-3 linkage to Gal^a
* have not shown terminations
- with sialic acid in alpha-2 -6 linkage to Gal^a
- with intersecting GIcNAc (GIcNAc-beta 1-4Man beta 1-4
- with Galc-alpha1-3Gal^b
- Via GalNAc-beta 1-4GlcNAc (N,N'-diacetyllactosediamine
type)^c
^a NeuSAc in alpha2-6 linkage has only been reported by
Davidson and Castellino (84).
^b This termination ha. only be described by Ashford, et al.
(85).
^c A GalNAc-transferase activity has been detected in CHO
cells by Dharmesh, et al. (86).
--------------------------------------------------------------
So far, studies have mainly been reported, describing the generation of recombinant human EPO in cells having a glycosylation machinery, such as CHO cells and baby hamster kidney (BHK) cells. With respect to the primary structure analysis of the carbohydrate chains of urinary EPO and recombinant EPO expressed in CHO and BHK cells, several investigations have been published (Urinary EPO: see 106-109; CHO EPO: 43, 106, 108-114; BHK EPO: 107, 115-117). It should be emphasized that the various research groups studied different batches of EPO's supplied by different companies. Also the use of other cell lines has been evaluated, and typical examples comprise psi 2 cells derived from NIH/3T3 (118), insect cells (119), and cultured tobacco cells (120).
--------------------------------------------------------------
Table 4. Monosaccharide analysis data of urinary and CHO
erythropoietins (43, 106). Mannose is taken as 3.0.
-------------------------------------------------------------
Fuc Man Gal GlcNAc GalNAc Sialic
acid
-------------------------------------------------------------
Urinary 0.9 3.0 4.2 5.3 0.3 3.4
EPO
CHO EPO 0.8-1.4 3.0 4.3-5.7 5.9-6.5 trace-0.5 3.1-4.3
--------------------------------------------------------------
In Table 4, the monosaccharide analysis data of urinary and CHO EPO's, reported in the literature (43, 106), are compared (see also 107). At a first glance these data are quite similar; although a tendency may exist in that CHO EPO may contain slightly higher Gal and/or GlcNAc values. However, because different ensembles of carbohydrate chains can give rise to similar monosaccharide analyses, it is dangerous to draw detailed conclusions in terms of identity, similarity or non-identity of EPO's directly from such an analysis (see below).
The most detailed study so far on the N,O-glycosylation of EPO has been reported by Hokke, et al. (43, 49, 50) for CHO EPO. It includes the identification of over 35 different N- linked and two O-linked oligosaccharides, representing at least 95 mol/100 mol carbohydrate chains, and the results of this N- and O-glycosylation study are presented in Tables 5 and 6, respectively. Although the literature results of the various studies on the N,O-glycosylation of urinary, CHO and BHK EPO's are to some extent similar, discrepancies are observed on a more detailed level. The discrepancies hold even for probes of a similar biological origin, investigated by different research groups. It may be evident that the structural data published for EPO support the statement in the Introduction that primary structure analysis of glycoprotein glycans has still not reached the level of a routine analysis, which is also of importance for the growing interest in profiling techniques.
The studies reported by Tsuda, et al. (107) and Takeuchi, et al. (108) showed a clear tendency that CHO and BHK EPO's contain a higher content of N-linked carbohydrate chains with Galbeta1-4GlcNAcbeta1-3 (N- acetyllactosamine) repeats than the urinary form. In recombinant EPO's the repeating N-acetyllactosamine units were found to be, at any case, in the branches arising from the alpha1-6 linked Man residue: Gal-beta1-4GlcNAc-beta1- 3Gal-beta1-4GlcNAc-beta1-6Man-alpha1-6 and/or Gal-beta1- 4GlcNAc-beta1-3Gal-beta1-4GlcNAc-beta1-2Man-alpha1-6 (the only extensions reported by Hokke, et al. (43), Takeuchi, et al (108), Rice, et al. (109). For a survey of the N-acetyllactosamine extensions, as found by Hokke, et al. (43), see table 5.Focusing on tetraantennary glycans of CHO EPO also the sequence Gal- beta1-4GlcNAc-beta1-3Gal-beta1-4GlcNAc-beta1-4Man-alpha1-3 has been identified (106, 110). Small amounts of tetraantennary carbohydrate chains with three repeats have been reported in Sasaki, et al. (106), Watson, et al. (113) and Linsley, et al. (114). In this case the third extension can comprise the Galbeta1-4GlcNAc-beta1-3Gal-beta1-4GlcNAc- beta1-4Man-alpha1-3 element, but also the Gal-beta1-4GlcNAc- beta1-3Gal-beta1-4GlcNAc-beta1-2Man-alpha1-3 sequence has been identified (106). Also the occurrence of trimeric N-acetyllactosamine units has been mentioned to occur (106). Focusing on tetraantennary glycans of BHK EPO, also extensions with Gal-beta1-4GlcNAc-beta1-3Gal-beta1-4GlcNAc- beta1-2Man-alpha1-3 have been established (107, 116). The presence of repeats in di- and tri'antennary glycans of CHO EPO has been described by Hokke, et al. (43), and the presence of a repeat, Gal-beta1-4GlcNAc-beta1-3Gal-beta1- 4GlcNAc-beta1-4Man-alpha1-3, in triantennary glycans of CHO EPO by Sasaki, et al. (106) (see also 116). Linsley, et al. (114) reported the existence of repeats for both the tri- and the tri'antennary structures. For urinary and BHK EPO, the occurrence of a Gal-beta1-3GlcNAc-beta1-4Man-alpha1-3 instead of a Gal-beta1-4GlcNAc-beta1-4Man-alpha1-3 sequence in tri- and tetraantennary glycans has been proposed by Tsuda, et al. (107). In a few BHK EPO batches small amounts of N,N'-diacetyllactosediamine or hybrid type of chains were observed (116).
In all studies focused on CHO EPO's the linkage between sialic acid and galactose in the N-glycans was demonstrated to be exclusively alpha2-3 (Table 5), whereby Hokke, et al. (43, 49) showed that besides Neu5Ac as the major sialic acid (95 %) also Neu5Gc (2 %) and Neu5,9Ac2 (3 %) do occur. Linsley, et al. (114) described for each detected Neu5Ac-containing compound a satellite wherein one Neu5Ac residue was replaced by one Neu5Gc residue (see also Table 5). The same type of linkage with Neu5Ac was also established for BHK EPO's, and in this case Neu5Gc has been mentioned as a possible minor sialic acid constituent (116). Inconsistencies exist with respect to the type of linkage in urinary EPO glycan chains. Based on methylation analysis data, Sasaki, et al. (106) found evidence for the exclusive existence of alpha2-3 linkages, whereas Takeuchi, et al. (108) demonstrated the occurrence of both alpha2-3 (major) and alpha2-6 (occasionally) linkages. The occurrence of sulphate groups in urinary and CHO EPO has been suggested (108, 121), but structural studies have not been published thus far. With respect to the alpha1-6 fucosylation of the Asn-bound GlcNAc residues in CHO and BHK EPO's, amounts between at least 80 % (106, 108, 110) and nearly 100 % (43, 116) have been reported. In BHK EPO the presence of a mannose-6-phosphate group has been demonstrated (117).
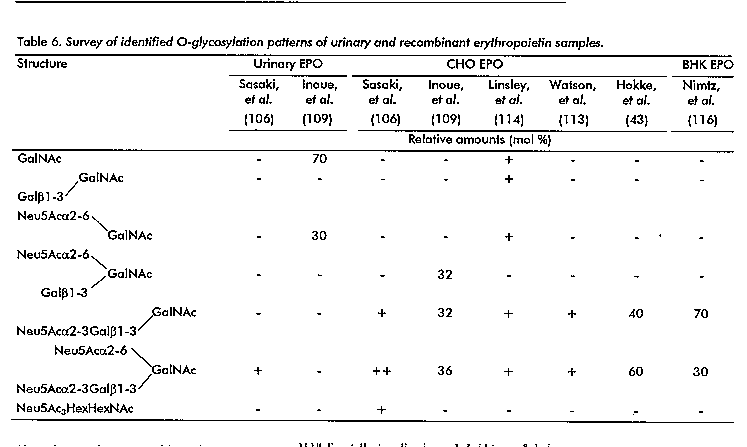
In Table 6 a survey of identified O-glycosylation patterns of urinary, CHO and BHK EPO's is presented. Comparing the results of the seven studies on recombinant EPO's (43, 106, 109, 110, 114-116) it is clear that the major carbohydrate chains are the trisaccharide Neu5Ac-alpha2-3Gal-beta1-3GalNAc and the tetrasaccharide Neu5Ac-alpha2-3Gal-beta1-3(Neu5Ac-alpha2-6) GalNAc. Additional O-linked chains comprise in general desialylated forms of these oligosaccharides. Interestingly, Linsley, et al. (114) also reported the disaccharide Neu5Acalpha2-6GalNAc and its desialylated form, the same saccharides as found by Inoue, et al. (109) in urinary EPO, and claimed to be unique for the urinary EPO. In the case of BHK EPO, Nimtz, et al. (116) published an O- glycosylation percentage of only 60 %. A similar finding of partial O-glycosylation was also reported for CHO EPO by Linsley, et al. (114).
The variations observed in the degree of sialylation, as found in the literature for recombinant EPO's, may have been caused by the use of different purification methods to obtain recombinant EPO (distribution of glycoforms which make up the final product). Other causes for the variation in the degree of sialylation and fucosylation could be the use of various cell clones, different culturing conditions, which can lead to differences in the activities of the alpha2-3 sialyltransferase or alpha1-6 fucosyltransferase, and the presence of sialidase activity for CHO cells recently reported in the culture fluid (122). In principle, the inconsistencies in sialylation may also have been caused partly by the use of different methods for the release and the purification of the carbohydrate chains. It should be noted that the non-random distribution of sialic acids in tetra- and diantennary glycans, as found by Hokke, et al. (43), is in good agreement with the branch specificity of human placental alpha2-3 sialyltransferase (123). With respect to the differences observed in the relative amounts of triantennary and tri'antennary oligosaccharides in various recombinant probes, it should be stated that these different ratios cannot be artefacts caused by analytical problems. They may be explained by differences in the activities of the involved glycosyltransferases for the various cell lines and culture conditions.
Finally, the site-specific N-glycosylation in CHO EPO has been studied by Sasaki, et al. (110) and Linsley, et al. (114). Di-, tri/tri'- and tetraantennary (major) glycans were detected for Asn-24, whereas Asn-38 and Asn-83 contained mainly tetraantennary chains. With respect to the site-specific N-glycosylation pattern in BHK EPO, it was established that Asn-24 was glycosylated with di-, tri/tri'- and tetraantennary chains, and hybrid-type chains, whereas Asn-38 and Asn-83 contained almost exclusively the tetraantennary chains (116). In addition, recently, a minor phosphorylated oligomannose-type carbohydrate chain, GlcNAc- alpha1-P-6Man-alpha1-2Man-alpha1-3[Manalpha1-6(Man- alpha1-3)Man-alpha1-6] Man-beta1-4GlcNAc-beta1-4GlcNAc, was found at Asn-24 (117). Acknowledgements The author gratefully acknowledge the contributions of Drs. AA Bergwerff, JBL Damm, P De Waard, GJ Gerwig, K Hard, CH Hokke, JHGM Mutsaers, and JFG Vliegenthart from the Bijvoet Center for Biomolecular Research at Utrecht University for their contributions to the different topics presented in this paper. These investigations were supported by the Netherlands Foundation for Chemical Research (SON) with financial aid from the Netherlands Organization for Scientifc Research (NWO), and the Netherlands Program for Innovation Oriented Carbohydrate Research (IOP-k) with financial aid from the Ministry of Economic Affairs and the Ministry of Agriculture, Nature Management and Fisheries. References 1 Rademacher TW, Parekh RB, Dwek RA. Glycobiology. Annu Rev Biochem 1989; 57: 785-838. 2 Allen HJ Kisailus EC (Eds.). Glycoconjugates - Composition, Structure and Function. Marcel Dekker, New York 1992. 3 Lis H, Sharon N. Protein glycosylation; structural and functional aspects. Eur J Biochem 1993;218:1-27. 4 Montreuil J, Vliegenthart JFG, Schachter H (Eds.). Glycoproteins. New Comprehensive Biochemistry. Elsevier Science Amsterdam 1995;29a. 5 Montreuil J, Vliegenthart JFG, SchachterH (Eds.). Glycoproteins. New Comprehensive Biochemistry Elsevier. Science Amsterdam 1996;29b. 6 Parekh RB, Dwek RA, Edge CJ, Rademacher TW. N-glycosylation and the production of recombinant glycoproteins. TIBTECH 1989;7:117-122 7 Paulson JC. Glycoproteins: what are the sugar chains for? TIBS 1989;14:272-276. 8 Goochee CF, Gramer MJ, Andersen DC, Bahr JB, Rasmussen JR. The oligosaccharides of glycoproteins:bioprocess factors affecting oligosaccharide structure and their effect on glycoprotein properties. Bio/Technology 1991;9:1347-1355. 9 Vliegenthart JFG, Dorland L, Van Halbeek H. High-resolution, ^1H-nuclear magnetic resonance spectroscopy as a tool in the structural analysis of carbohydrates related to glycoproteins. Adv Carbohydr Chem Biochem 1983;41:209-374. 10 Dell A FAB-Mass spectrometry of carbohydrates. Adv Carbohydr Chem Biochem 1987;45:19-72. 11 Kamerling JP, Vliegenthart JFG. High-resolution ^1H-nuclear magnetic resonance spectroscopy of oligosaccharide-alditols released from mucin-type O-glycoproteins. Biol Magn Reson 1992;10:1-194. 12 Kamerling JP, Vliegenthart JFG. Carbohydrates. In:Clinical Biochemistry - Principles, Methods, Applications, Vol. 1, Mass Spectrometry (LAWSON AM, Ed.), Walter de Gruyter, Berlin 1989:175-263. 13 Mccloskey JA (Ed.). Mass spectrometry. Methods Enzymol 1990;193. 14 Besada V, Antuch W, Rodriguez R, Gonzalez L, Chinea G, Padron G. Mass spectrometry in protein analysis. Biotecnologia Aplicada 1992;9:1-15. 15 Nguyen DN, Becker GW, Riggin RM. Protein mass spectrometry:applications to analytical biotechnology. J Chromatogr 1995;705:21-45. 16 Settineri CA, Burlingame AL. Mass spectrometry of carbohydrates and glycoconjugates. In: Carbohydrate Analysis - High Performance Liquid Chromatography and Capillary Electrophoresis (EL RASSI Z, Ed.), Elsevier Science, Amsterdam 1995: 447-514. 17 Fukuda M, A Kobata (Eds.). Glycobiology - A Practical Approach. IRL Press at Oxford University Press, Oxford 1993. 18 Hounsell EF (Ed.). Glycoprotein Analysis in Biomedicine. Meth Mol Biol, Vol. 14. Humana Press Inc., Totowa 1993. 19 Lennarz WJ, Hart GW (Eds.). Guide to Techniques in Glycobiology. Methods Enzymol 1994;230. 20 Dwek RA, Edge CJ, Harvey DJ, Wormald MR, Parekh RB. Analysis of glycoprotein-associated oligosaccharides. Annu Rev Biochem 1993;62:65-100. 21 Verbert A (Ed.). Methods on Glycoconjugates. Harwood Academic Publishers, Reading 1995. 22 Yamashita K, Mizuochi T, Kobata A. Analysis of oligosaccharides by gel filtration. Methods Enzymol 1982;83:105-126. 23 Hard K, Van Zadelhoff G, Moonen P, Kamerling JP, Vliegenthart JFG. The Asn-linked carbohydrate chains of human Tamm-Horsfall glycoprotein of one male; novel sulfated and novel N- acetylgalactosamine-containing N-linked carbohydrate chains. Eur J Biochem 1992;209:895-915. 24 Hokke CH, Damm JBL, Penninkhof B, Aitken RJ, Kamerling JP, Vliegenthart JFG. Structure of the O-linked carbohydrate chains of porcine zona pellucida glycoproteins. Eur J Biochem 1994;221:491-512. 25 Tomiya N, Awaya J, Kurono M, Endo S, Arata Y, Takahashi N. Analyses of N-linked oligosaccharides using a two-dimensional mapping technique. Anal Biochem 1988;171:73-90. 26 Hermentin P, Witzel R, Vliegenthart JFG, Kamerling JP, Nimtz M, Conradt HS. A strategy for the mapping of N-glycans by high-pH anion-exchange chromatography with pulsed amperometric detection. Anal Biochem 1992;203:281-289. 27 Townsend RR. Analysis of glycoconjugates using high-pH anion- exchange chromatography. In: Carbohydrate Analysis -High Performance Liquid Chromatography and Capillary Electrophoresis (EL RASSI Z, Ed.), Elsevier Science, Amsterdam 1995: 181-209. 28 Hermentin P, Doenges R, Witzel R, Hokke CH, Vliegenthart JFG, Kamerling JP, Conradt HS, Nimtz M, Brazel D. A strategy for the mapping of N-glycans by high-performance capillary electrophoresis. Anal Biochem 1994;221:29-41. 29 Suzuki S, Honda S. Two-dimensional mapping of N-glycosidically linked oligosaccharides in glycoproteins by high-performance capillary electrophoresis. TRAC 1995; 14:279-288. 30 Hu G-F. Fluorophore-assisted carbohydrate electrophoresis: technology and applications. J Chromatogr 1995; 705:89-103. 31 Edge CJ, Rademacher TW, Wormald MR, Parekh RB, Butters TD, Wing DR, Dwek RA. Fast sequencing of oligosaccharides: the reagent-array analysis method. Proc Nat Acad Sci USA 1992;89:6338-6342. 32 Bergwerff AA, Van Oostrum J, Asselbergs Fam, Burgi R, Hokke CH, Kamerling JP, Vliegenthart JFG. Primary structure of N- linked carbohydrate chains of a human chimeric plasminogen activator K2tu-PA expressed in Chinese hamster ovary cells. Eur J Biochem 1993;212:639-656. 33 Bergwerff AA, Van Dam GJ, Rotmans JP, Deelder AM, Kamerling JP, Vliegenthart JFG. The immunologically reactive part of immunopurified circulating anodic antigen from Schistosoma mansoni is a threonine-linked polysaccharide consisting of -->6)-beta-D-GlcpA-(1-->3))-beta-D-GalpNAc-(1--> repeating units. J Biol Chem 1994;269: 31510-31517. 34 Bergwerff AA, Stroop CJM, Murray B, -P Holtorf A, Pluschke G, Van Oostrum J, Kamerling JP, Vliegenthart JFG. Variation in N- linked carbohydrate chains in different batches of two chimeric monoclonal IgG1 antibodies produced by different murine SP2/0 transfectoma cell subclones. Glycoconjugate J 1995;12:318-330. 35 Bergwerff AA, Van Oostrum J, Kamerling JP, Vliegenthart JFG. The major N-linked carbohydrate chains from human urokinase; the occurrence of 4-O-sulfated, (alpha2-6)-sialylated or (alpha1-3)-fucosylated N-acetylgalactosamine (beta1- 4)-N-acetylglucosa-mine elements. Eur J Biochem 1995b; 228: 1009-1019. 36 Damm JBL, Hard K, Kamerling JP, Van Dedem GWK, Vliegenthart JFG. Structure determination of the major N- and O-linked carbohydrate chains of the beta subunit from equine chorionic gonadotropin. Eur J Biochem 1990;189:175-183. 37 De Waard P, Koorevaar A, Kamerling JP, Vliegenthart JFG. Structure determination by ^1H NMR spectroscopy of (sulfated) sialylated N-linked carbohydrate chains released from porcine thyroglobulin by peptide-N4-(N-acetyl-beta- glucosaminyl) asparagine amidase-F. J Biol Chem 1991; 266:4237-4243. 38 Gerwig GJ, Kamerling JP, Vliegenthart JFG, Morag E, Lamed R, Bayer EA. The nature of the carbohydrate-peptide linkage region in glycoproteins from the cellulosomes of Clostridium thermocellum and Bacteroides cellulosolvens. J Biol Chem 1993; 268:26956-26960. 39 Hard K, Mekking A, Damm JBL, Kamerling JP, De Boer W, Wijnands RA, Vliegenthart JFG. Isolation and structure determination of the intact sialylated N-linked carbohydrate chains of recombinant human follitropin expressed in Chinese hamster ovary cells. Eur J Biochem 1990; 193:263-271. 40 Hard K, Damm JBL, Spruijt MPN, Bergwerff AA, Kamerling JP, Van Dedem GWK, Vliegenthart JFG. The carbohydrate chains of the b subunit of human chorionic gonadotropin produced by the choriocarcinoma cell line BeWo; novel O-linked and novel bisecting-GlcNAc-containing N-linked carbohydrates. Eur J Biochem 1992; 205: 785-798.
41 Hard K, Van Doorn JM, Thomas-Oates JE, Kamerling JP, Van Der Horst DJ. Structure of the Asn-linked oligosaccharides of apolipophorin III from the insect Locusta migratoria; carbohydrate-linked 2-aminoethylphosphonate as a constituent of a glycoprotein. Biochemistry 1993;32:766-775. 42 Hokke CH, Roosenboom MJH, Thomas-Oates JE, Kamerling JP, Vliegenthart JFG. Structure determination of the disialylated poly-(N-acetyllactosamine)-containing O-linked carbohydrate chains of equine chorionic gonadotropin. Glycoconjugate J 1994; 11:35-41. 43 Hokke CH, Bergwerff AA, Van Dedem GWK, Kamerling JP, Vliegenthart JFG. Structural analysis of the sialylated N- and O-linked carbohydrate chains of recombinant human erythropoietin expressed in Chinese hamster ovary cells; sialylation patterns and branch location of dimeric N- acetyllactosamine units. Eur J Biochem 1995;228:981-1008. 44 Kubelka V, Altmann F, Staudacher E, Tretter V, Marz L, Hard K, Kamerling JP, Vliegenthart JFG. Primary structures of the N-linked carbohydrate chains from honeybee venom phospholipase A2. Eur J Biochem 1993;213:1193-1204. 45 Pfeiffer G, Stirm S, Geyer R, Strube K-H, Bergwerff AA, Kamerling JP, Vliegenthart JFG. Structure elucidation of sulphated oligosaccharides from recombinant human tissue plasminogen activator expressed in mouse epithelial cells. Glycobiology 1992; 2:411-418. 46 Van Dam GJ, Bergwerff AA, Thomas-Oates JE, Rotmans JP, Kamerling JP, Vliegenthart JFG, Deelder AM. The immunologically reactive O-linked polysaccharide chains derived from circulating cathodic antigen isolated from the human blood fluke Schistosoma mansoni have Lewis x as repeating unit. Eur J Biochem 1994 225: 467-482. 47 Van Zuylen Cwem, De Beer T, Rademaker GJ, Haverkamp J, Thomas- Oates JE, K Hard, Kamerling JP, Vliegenthart JFG. Site- specific and complete enzymic deglycosylation of the native human chorionic gonadotropin a-subunit. Eur J Biochem 1995; 231:754-760. 48 Mutsaers JHGM, Kamerling JP, Devos R, Guisez Y, Fiers W, Vliegenthart JFG. Structural studies of the carbohydrate chains of human g-interferon. Eur J Biochem 1986; 156:651- 654. 49 Hokke CH, Bergwerff AA, Van Dedem GWK, Van Oostrum J, Kamerling JP, Vliegenthart JFG. Sialylated carbohydrate chains of recombinant human glycoproteins expressed in Chinese hamster ovary cells contain traces of N-glycolylneuraminic acid. FEBS Lett 1990;275: 9-14. 50 Hokke CH, Kamerling JP, Van Dedem GWK, Vliegenthart JFG. Determination of the branch location of extra N- acetyllactosamine units in sialo N-linked tetraantennary oligosaccharides. FEBS Lett 1991; 286:18-24. 51 Hard K, Bitter W, Kamerling JP, Vliegenthart JFG. O- Mannosylation of recombinant human insulin-like growth factor I (IGF-I) produced in Saccharomyces cerevisiae. FEBS Lett 1989;248:111-114. 52 Dan K, Andreasen PA, Gr ndahl-Hansen J, Kristensen P, Nielsen LS, Skriver L. Plasminogen activators, tissue degradation, and cancer. Adv Cancer Res 1985;44:139-266. 53 Wun T-C. Plasminogen activation: biochemistry, physiology, and therapeutics. CRC Crit Rev Biotech 1988;8:131-148. 54 Lijnen HR, Collen D. Strategies for the improvement of thrombolytic agents. Thromb Haemostas 1991;66:88-110. 55 Hansen L, Blue Y, Barone K, Collen D, Larsen GR. Functional effects of asparagine-linked oligosaccharide on natural and variant human tissue-type plasminogen activator. J Biol Chem 1988; 263:15713-15719. 56 Beebe DP, Aronson DL. Turnover of tPA in rabbits: influence of carbohydrate moieties. Thromb Res 1988;51:11-22. 57 Collen D, M Stassen J, Larsen G. Pharmacokinetics and thrombolytic properties of deletion mutants of human tissue- type plasminogen activator in rabbits. Blood 1988;71:216- 219. 58 Lucore CL, Fry Eta, Nachowiak DA, Slobe BE. Biochemical determinants of clearance of tissue-type plasminogen activator from the circulation. Circulation 1988; 77:906-914. 59 Wittwer AJ, Howard SC, Carr LS, Harakas NK, Feder J, Parekh RB, Rudd PM, Dwek RA, Rademacher TW. Effects of N- glycosylation on in vitro activity of Bowes melanoma and human colon fibroblast derived tissue plasminogen activator. Biochemistry 1989;28:7662-7669. 60 Wittwer AJ, Howard SC. Glycosylation at Asn-184 inhibits the conversion of single-chain to two-chain tissue-type plasminogen activator by plasmin. Biochemistry 1990; 29:4175- 4180. 61 Howard SC, Wittwer AJ, WelplY JK. Oligosaccharides at each glycosylation site make structure-dependent contributions to biological properties of human tissue plasminogen activator. Glycobiology 1991;1: 411-417. 62 Otter M, Barrett-Bergshoeff MM, Rijken DC. Binding of tissue- type plasminogen activator by the mannose receptor. J Biol Chem 1991;266:13931-13935. 63 Berg DT, Burck PJ, Berg DH, Grinnell BW. Kringle glycosylation in a modified human tissue plasminogen activator improves functional properties. Blood 1993;81:1312-1322. 64 Hajjar KA, Reynolds CM. alpha-Fucose-mediated binding and degradation of tissue-type plasminogen activator by HepG2 cells. J Clin Invest 1994;93:703-710. 65 Parekh RB, Dwek RA, Thomas JR, Opdenakker G, Rademacher TW, Wittwer AJ, Howard SC, Nelson R, Siegel NR, Jennings MG, Harakas NK, Feder J. Cell-type-specific and site-specific N- glycosylation of type I and type II human tissue plasminogen activator. Biochemistry 1989;28:7644-7662. 66 Pohl G, Kenne L, Nilsson B, Einarsson M. Isolation and characterization of three different carbohydrate chains from melanoma tissue plasminogen activator. Eur J Biochem 1987;170:69-75. 67 Chan AL, Morris HR, Panico M, Etienne AT, Rogers ME, Gaffney P, Creighton-Kempsford L, Dell A. A novel sialylated N- acetylgalactosamine-containing oligosaccharide is the major complex-type structure present in Bowes melanoma tissue plasminogen activator. Glycobiology 1991; 1:173-185. 68 Carr SA, Roberts GD, Jurewicz A, Frederick B. Structural fingerprinting of Asn-linked carbohydrates from specific attachment sites in glycoproteins by mass spectrometry: application to tissue plasminogen activator. Biochimie 1988;70:1445-1454. 69 Parekh RB, Dwek RA, Rudd PM, Thomas JR, Rademacher TW, Warren T, Wun T-C, Hebert B, Reitz B, Palmier M, Ramabhadran T, Tiemeier DC. N-glycosylation and in vitro enzymatic activity of human recombinant tissue plasminogen activator expressed in Chinese hamster ovary cells and a murine cell line. Biochemistry 1989;28:7670-7679. 70 Spellman MW, Basa LJ, Leonard CK, Chakel JA, O'Connor JV, Wilson S, Van Halbeek H. Carbohydrate structures of human tissue plasminogen activator expressed in Chinese hamster ovary cells. J Biol Chem 1989;264:14100-14111. 71 Pfeiffer G, Schmidt M, Strube K-H, Geyer R. Carbohydrate structure of recombinant human uterine tissue plasminogen activator expressed in mouse epithelial cells. Eur J Biochem 1989;186:273-286. 72 Nimtz M G Noll, Paques E-P, Conradt HS. Carbohydrate structures of a human tissue plasminogen activator variant expressed in recombinant Chinese hamster ovary cells. FEBS Lett 1990;271:14-18. 73 Pfeiffer G, Strube K-H, Schmidt M, Geyer R. Glycosylation of two recombinant human uterine tissue plasminogen activator variants carrying an additional N-glycosylation site in the epidermal-growth-factor-like-domain. Eur J Biochem 1994; 219:331-348. 74 Wilhelm J, Lee SG, Kalyan NK, Cheng SM, Wiener F, Pierzchala W, Hung PP. Alterations in the domain structure of tissue-type plasminogen activator change the nature of asparagine glycosylation. Bio/Tech-nology 1990;8:321-325. 75 Minch SL, Kallio PT, Bailey JE. Tissue plasminogen activator coexpressed in Chinese hamster ovary cells with alpha(2,6)- sialyltransferase contains NeuAcalpha(2,6) Galbeta(1,4) GlcNAcR linkages. Biotechnol Progress 1995; 11:348-351. 76 Schmitt M, Janicke F, Moniwa N, Chucholowski N, Pache L, Graeff H. Tumor-associated urokinase-type plasminogen activator:biological and clinical significance. Biol Chem Hoppe-Seyler 1992; 373:611-622. 77 Zamarron C, Lijnen HR, Van Hoef B, Collen D. Biological and thrombolytic properties of proenzyme and active forms of human urokinase. I. Fibrinolytic and fibrinogenolytic properties in human plasma in vitro of urokinases obtained from human urine or by recombinant DNA technology. Thromb Haemostas 1984;52:19-23. 78 Sarubbi E, Nolli ML, Robbiati F, Soffientini A, Parenti F, Cassani G. The differential glycosylation of human pro-urokinase from various recombinant mammalian cell lines does not affect activity and binding to PAI-1. Thromb Haemostas 1989;62:927-933. 79 Henkin J, Dudlak D, Beebe DP, Sennello L. High sialic acid content slows prourokinase turnover in rabbits. Thromb Res 1991; 63:215-225. 80 Li X-K, Lijnen HR, Nelles L, Hu M-H, Collen D. Biochemical properties of recombinant mutants of nonglycosylated single chain urokinase-type plasminogen activator. Biochim Biophys Acta 1992; 1159:37-43. 81 Lenich C, Pannell R, Henkin J, Gurewich V. The influence of glycosylation on the catalytic and fibrinolytic properties of pro-urokinase. Thromb Haemostas 1992; 68: 539-544. 82 Bergwerff AA, Thomas-Oates JE, Van Oostrum J, Kamerling JP, Vliegenthart JFG. Human urokinase contains GalNAcbeta(1- 4)[Fucalpha(1-3)]GlcNAcbeta(1-2) as a novel terminal element in N-linked carbohydrate chains. FEBS Lett 1992;314:389- 394. 83 Asselbergs FAM, Burgi R, Chaudhuri B, Heim J, Meyhack B, Rajput B, Vna Oostrum J, Alkan S. Localization of epitopes by monoclonal antibodies on tissue-type and urokinase-type plasminogen activators using recombinant hybrid enzymes. Fibrinolysis 1993;7:1-14. 84 Davidson DJ, Castellino FJ. Oligosaccharide structures present on asparagine-289 of recombinant human plasminogen expressd in a Chinese hamster ovary cell line. Biochemistry 1991;30:625- 633. 85 Ashford DA, Alafi CD, Gamble VM, Mackay DJG, Rademacher TW, Williams PJ, Dwek RA, Barclay AN, Davis SJ, Somoza C, Ward HA, Williams AF. Site-specific glycosylation of recombinant rat and human soluble CD4 variants expressed in Chinese hamster ovary cells J Biol Chem 1993; 268:3260-3267. 86 Dharmesh SM, Skelton TP, Baenziger JU. Co-ordinate and restricted expression of the ProXaaArg/Lys-specific GalNAc- transferase and the GalNAcb1, 4GlcNAcb1, 2Mana-4- sulfotransferase. J Biol Chem 1993; 268:17096-17102. 87 Ashwell G, Harford J. Carbohydrate-specific receptors of the liver. Annu Rev Biochem 1982;51:531-554. 88 Fiete D, Srivastava V, Hindsgaul O, Baenziger JU. A hepatic reticuloendothelial cell receptor specific for SO4^-- 4GalNAcbeta1,4GlcNAcbeta1,2Manalpha that mediates rapid clearance of lutropin Cell 1991; 67:1103-1110. 89 Drickamer K Clearing up glycoprotein hormones. Cell 1991;67:1029-1032. 90 Chiu MH, Tamura T, Wadhwa MS, Rice KG. In vivo targetting function of N-linked oligosaccharides with terminating galactose and N-acetylgalactosamine residues. J Biol Chem 1994;269:16195-16202. 91 Krantz SB. Erythropoietin. Blood 1991; 77:419-434. 92 Lai P-H, Everett R, Wang F-F, Arakawa T, Goldwasser E. Structural characterization of human erythropoietin. J Biol Chem 1986; 261:3116-3121. 93 Koury MJ, Bondurant MC. The molecular mechanism of erythropoietin action. Eur J Biochem 1992;210:649-663. 94 Roberts D, Smith DJ. Erythropoietin: induction of synthesis to signal transduction. J Mol Endocrinol 1994;12:131-148. 95 Takeuchi M, Kobata A. Structures and functional roles of the sugar chains of human erythropoietins. Glycobiology 1991; 1:337-346. 96 Storring, PL, Gaines Das RE. The international standard for recombinant DNA-derived erythropoietin:collaborative study of four recombinant DNA-derived erythropoietins and two highly purified human urinary erythropoietins. J Endocrinol 1992; 134:459-484. 97 Dube S, Fisher JW, Powell JS. Glycosylation at specific sites of erythropoietin is essential for biosynthesis, secretion, and biological function. J Biol Chem 1988;263: 17516-17521. 98 Fukuda MN, Sasaki H, Lopez L, Fukuda M. Survival of recombinant erythropoietin in the circulation:the role of carbohydrates. Blood 1989;73:84-89. 99 Imai N, Higuchi M, Kawamura A, Tomonoh K, Oh-Eda M, Fujiwara M, Shimonaka Y, Ochi N. Physicochemical and biological characterization of asialoerythropoietin; suppressive effects of sialic acid in the expression of biological activity of human erythropoietin in vitro. Eur J Biochem 1990;194:457- 462. 100 Takeuchi M, Takasaki S, Shimada M, Kobata A. Role of sugar chains in the in vitro biological activity of human erythropoietin produced in recombinant Chinese hamster ovary cells. J Biol Chem 1990; 265:12127-12130. 101 Owers Narhi L, Arakawa T, Aoki KH, Elmore R, Rohde MF, Boone T, Strickland TW. The effect of carbohydrate on the structure and stability of erythropoietin. J Biol Chem 1991;266:23022- 23026. 102 Sytkowski AJ, Feldman L, Zurbuch DJ. Biological activity and structural stability of N-deglycosylated recombinant human erythropoietin. Biochem Biophys Res Commun 1991;176:698- 704. 103 Delorme E, Lorenzini T, Giffin J, Martin F, Jacobsen F, Boone T, Elliott S. Role of glycosylation on the secretion and biological activity of erythropoietin. Biochemistry 1992; 31:9871-9876. 104 Yamaguchi K, Akai K, Kawanishi G, Ueda M, Masuda S, Sasaki R. Effects of site-directed removal of N-glycosylation sites in human erythropoietin on its production and biological properties. J Biol Chem 1991; 266:20434-20439. 105 Higuchi M, Oh-Eda M, Kuboniwa H, Tomonoh K, Shimonaka Y, Ochi N. Role of sugar chains in the expression of the biological activity of human erythropoietin. J Biol Chem 1992;267:7703-7709. 106 Sasaki H, Bothner B, Dell A, Fukuda M. Carbohydrate structure of erythropoietin expressed in Chinese hamster ovary cells by a human erythropoietin cDNA. J Biol Chem 1987;262:12059- 12076. 107 Tsuda E, Goto M, Murakami A, Akai K, Ueda M, Kawanishi G, Takahashi N, Sasaki R, Chiba H, Ishihara H, Mori M, Tejima S, Endo S, Arata Y. Comparative structural study of N-linked oligosaccharides of urinary and recombinant erythropoietins. Biochemistry 1988;27:5646-5654. 108 Takeuchi M, Takasaki S, Miyazaki H, Kato T, Hoshi S, Kochibe N, Kobata A. Comparative study of the asparagine-linked sugar chains of human erythropoietin purified from urine and the culture medium of recombinant Chinese hamster ovary cells. J Biol Chem 1988;263:3657-3663. 109 Inoue N, Takeuchi M, Asano K, Shimizu R, Takasaki S, Kobata A. Structures of mucin-type sugar chains on human ery- thropoietins purified from urine and the culture medium of recombinant Chinese hamster ovary cells. Arch Biochem Biophys 1993;301:375-378. 110 Sasaki H, Ochi N, Dell A, Fukuda M. Site-specific glycosylation of human recombinant erythropoietin:analysis of glycopeptides or peptides at each glycosylation site by fast atom bombardment mass spectrometry. Biochemistry 1988; 27:8618-8626. 111 Takeuchi M, Inoue N, Strickland TW, Kubota M, Wada M, Shimizu R, Hoshi S, KozutsumI H, Takasaki S, Kobata A. Relationship between sugar chain structure and biological activity of recombinant human erythropoietin produced in Chinese hamster ovary cells. Proc Natl Acad Sci USA 1989;86:7819-7822. 112 Rice Kg, Takahashi N, Namiki Y, Tran Ad, Lisi Pj, Lee Yc. Quantitative mapping of the N-linked sialyloligosaccharides of recombinant erythropoietin:combination of direct high- performance anion-exchange chro-matography and 2-aminopyridine derivatization. Anal Biochem 1992;206:278-287. 113 Watson E, Bhide A, Van Halbeek H. Structure determination of the intact major sialylated oligosaccharide chains of recombinant human erythropoietin expressed in Chinese hamster ovary cells. Glycobiology 1994;4:227-237. 114 Linsley KB, Chan S-Y, Chan S, Reinhold BB, Lisi PJ, Reinhold VN. Applications of electrospray mass spectrometry to erythropoietin N- and O-linked glycans. Anal Biochem 1994;219:207-217. 115 Tsuda E, Kawanishi G, Ueda M, Masu-da S, Sasaki R. The role of carbohydrate in recombinant erythropoietin. Eur J Biochem 1990;188:405-411. 116 Nimtz M, Martin W, Wray V, Kl”ppel K-D, Augustin J, ConradT HS. Structures of sialylated oligosaccharides of human erythropoietin expressed in recombinant BHK-21 cells. Eur J Biochem 1993; 213:39-56. 117 Nimtz M, Wray V, Rudiger A, Conradt HS. Identification and structural characterization of a mannose-6-phosphate containing oligomannosidic N-glycan from human erythropoietin secreted by recombinant BHK-21 cells. FEBS Lett 1995;365:203- 208. 118 Goto M, Akai K, Murakami A, Hashimoto C, Tsuda E, Ueda M, Kawanishi G, Takahashi N, Ishimoto A, Chiba H, Sasaki R. Production of recombinant human erythropoietin in mammalian cells:host-cell dependency of the biological activity of the cloned glycoprotein. Bio/Technology 1988; 6:67-71. 119 Wojchowski DM, Orkin SH, Sytkowski AJ. Active human erythropoietin expressed in insect cells using a baculovirus vector:a role for N-linked oligosaccharide. Biochim Biophys Acta 1987;910:224-232. 120 Matsumoto S, Ikura K, Ueda M, Sasaki R. Characterization of a human glycoprotein (erythropoietin) produced in cultured tobacco cells. Plant Mol Biol 1995; 27:1163-1172. 121 Strickland TW, Adler B, Aoki k, Asher S, Derby P, Goldwasser E, Rogers G. Occurrence of sulfate on the N-linked oligosaccharides of human erythropoietin. J Cell Biochem suppl 1992;16B:167. 122 Gramer MJ, Goochee CF, Chock VY, Brousseau DT, Sliwkowski MB. Removal of sialic acid from a glycoprotein in CHO cell culture supernatant by action of an extracellular CHO cell sialidase. Bio/Techno-logy 1995;13:692-698.
123 Nemansky M. Sialyltransferases:their specificity and their use in carbohydrate remodelling. Thesis Free University Amsterdam 1993. Copyright 1996 Elfos Scientiae
The following images related to this document are available:Line drawing images[ba96073h.gif] [ba96073a.gif] [ba96073d.gif] [ba96073b.gif] [ba96073e.gif] [ba96073g.gif] [ba96073c.gif] [ba96073f.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}